Download
1 / 1
10 likes | 123 Views
PERCEPTION OF THE STEROID MOLTING HORMONE (20-HE) MODIFIES AGGRESSIVE BEHAVIOR IN THE AMERICAN LOBSTER, Homarus americanus . Stuart Cromarty 1 , Debra Coglianese 2 Lenore Martin 3 and G. Kass-Simon 2

E N D
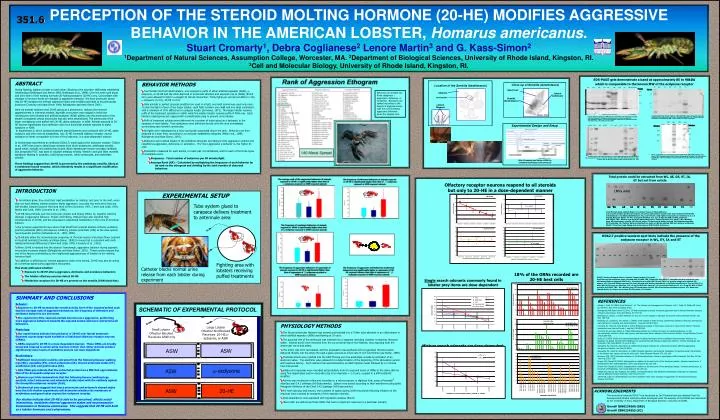
PERCEPTION OF THE STEROID MOLTING HORMONE (20-HE) MODIFIES AGGRESSIVE BEHAVIOR IN THE AMERICAN LOBSTER, Homarus americanus. Stuart Cromarty1, Debra Coglianese2 Lenore Martin3 and G. Kass-Simon2 1Department of Natural Sciences, Assumption College, Worcester, MA. 2Department of Biological Sciences, University of Rhode Island, Kingston, RI. 3Cell and Molecular Biology, University of Rhode Island, Kingston, RI. 351.6 SDS-PAGE gels demonstrate a band at approximately 85 to 98kDA which is comparable to the known MW of the ecdysone receptor ABSTRACT During fighting, lobsters urinate on each other. Blocking urine excretion obliterates established relationships (Breithaupt and Atema 1993, Breithaupt et al., 1999). Over the molt cycle blood and urine titers of the molting hormone 20-hydroxyecdysone (20-HE) vary. Concomitant with changes in hormone levels are changes in aggressive behavior. We have previously shown that 20-HE increases the internal aggressive state and modifies potentials at neuromuscular junctions (Cromarty and Kass-Simon 1998, Bolingbroke and Kass-Simon 2001). Here we present evidence that 20-HE acting as a pheromone, reduces the inherent aggressiveness in intermolt animals. Agonistic encounters were staged in which the nephropores were blocked and artificial seawater (ASW) puffed onto the antennules of the smaller combatants whose antennules had also been desensitized. The antennules of the larger combatants were puffed with 20-HE, alpha ecdysone, or ASW. Animals exposed to 20-HE became significantly more defensive and more timid than animals exposed to alpha ecdysone or ASW. In experiments in which isolated antennule lateral filaments were perfused with 20-HE, alpha ecdysone and other steroid metabolites, only 20-HE increased olfactory receptor neuron excitation to levels comparable to those of food odorants, in a dose-dependent manner. In biochemical experiments an antibody (DDA2.7) made against the ecdysone receptor (Talbot et al., 1993) was used to label tissue extracts from whole antennules, aesthetasc sensilla, guard setae, eyestalk and walking-leg muscle. Biotin-labeled pan-mouse secondary antibody and strepavidin-FITC was used to visualize antibody binding. Western and spot blots revealed significant labeling in eyestalks, walking-leg muscles, whole antennules, and aesthetasc sensilla. These findings suggest that 20-HE is perceived by the aesthetasc sensilla, likely at a membrane bound receptor, which ultimately results in a significant modification of aggressive behavior. • BEHAVIOR METHODS • Two female intermolt adult lobsters, one exposed to puffs of either artificial seawater (ASW), -ecdysone, or 20-HE and the other deprived of antennule olfaction and exposed only to (ASW) (5ml/5 min) were allowed to fight in a staged 30 minute interaction. Thirty fights are carried out ASW (n=10), -ecdysone (n=10), 20-HE (n=10). • Only animals in perfect physical condition are used in a fight, and each animal was used only once. To bias the fight in favor of the larger lobster, each fight contains one small and one large combatant, with a minimum of 10% difference in carapace length (Scrivener, 1971). The larger lobster receives puffs of the treatment substance in ASW, while the smaller lobster receives puffs of ASW only. Each lobster’s nephropores are capped with a polyethylene plug to prevent urine release. • Puffs of treatment solution were delivered via a system of tubes glued as a backpack to the carapace of each lobster. Test substances were delivered directly onto the area immediately surrounding each lobster’s antennules. • All fights were videotaped by a Sony camcorder suspended above the tank. Behaviors are then analyzed at a later time, according to a previously established ethogram (Mello et al., 1999; Bolingbroke and Kass-Simon, 2001). • Behaviors were ranked based on the published ethogram according to their aggressive content and classified as aggressive, defensive, or avoidance. The more aggressive a behavior is, the higher its rank. • Parameters measured for each animal, for each pair of combatants, and for each of the three types of treatments were: • Frequency - Total number of behaviors per 30 minute fight. • Average Rank (AR) – Calculated by multiplying the frequency of each behavior by its rank on the ethogram and dividing by the total number of observed behaviors. Behaviors are divided into three categories – aggressive, defensive, or avoidance. Behaviors are ranked according to the degree of aggressiveness, with the most intensely aggressive behaviors given the highest rank. Olfactory receptor neurons respond to all steroids but only to 20-HE in a dose-dependent manner • INTRODUCTION • As lobsters grow, they shed their rigid exoskeleton by molting. Just prior to the molt, when they are hard-shelled, lobsters become highly aggressive. Just after the molt when they are soft-shelled, lobsters become timid and tend to flee (Scrivener, 1971; Tamm and Cobb, 1978; Atema and Cobb, 1980; Cromarty et al., 1991). • 20-HE rises and falls over the molt-cycle (Snyder and Chang 1991a, b), together with the changes in aggressive behavior. Snyder and Chang, 1991a,b have also reported high concentrations of 20-HE, and the subsequent ecdysteroid metabolites in the urine of American lobsters. • Our previous experiments have shown that blood from premolt lobsters enhance excitatory junction potentials (EJPs) and depress inhibitory junction potentials (IJPs) at the claw-opener neuromuscular junction (Schwanke et al., 1983, 1990). • 20-HE also alters the neuromuscular properties of the claw-opener and phasic flexor systems in intermolt animals (Cromarty and Kass-Simon, 1998) in a way that is consistent with molt-related behavioral differences (Tamm and Cobb, 1978; Cromarty et al., 1991). • When 20-HE is injected into the lobsters' hemolymph, aggressive behavior during agonistic encounters increases sharply (Bolingbroke and Kass-Simon, 2001). These results indicate that one of the factors contributing to the heightened aggressiveness of lobsters is the molting hormone itself. • In addition to affecting the internal aggressive state of an animal, 20-HE may also be acting as a chemical signal during aggressive encounters. • This study addresses whether: • Exposure to 20-HE alters aggressive, defensive and avoidance behaviors • The lobster olfactory neurons detect 20-HE • Membrane receptors for 20-HE are present on the sensilla (ORN dendrites) EXPERIMENTAL SETUP Tube system glued to carapace delivers treatment to antennule area DDA2.7 positive western spot blots indicate the presence of the ecdysone receptor in WL, EY, IA and ST Fighting area with lobsters receiving puffed treatments Catheter blocks normal urine release from each lobster during experiment 18% of the ORNs recorded are 20-HE best cells DDA2.7 Positive Western Blots on Lobster Tissue Extracts (1uL each).Spot Blots on Nitrocellulose paper, blocked with 5%BSA-TBS, Primary Ab was DDA2.7, secondary Ab was polyclonal goat antimouse1IgGs conjugated with biotin. Visualization was with HRP-SA and TMB. 1uL protein sample was spotted per square, all 12 top and bottom squares (24 total) labeled with Braille indentations to facilitate identification. Top row left (#1) and right (#12) spots are identical 1 uL x 5 mg/mL BSA std. Spots #2-#11 are serial dilutions of 1 uL x 5 mg/mL BSA x 100uL previous std/100uL TBS. WL=Walking legs, AE=Aesthetasc sensilla, GS=Guard setae mechanoreceptors, ST= Antennule shaved Stalks, IA=Intact antennules, EY=Eyestalks, CU=Cuticle. Single search odorants commonly found in lobster prey items are dose-dependent • SUMMARY AND CONCLUSIONS • Behavior: • Exposure to 20-HE increases the overall activity level of the exposed animal such that the average rank of aggressive behaviors, the frequency of defensive and avoidance behaviors are increased. • The opponents of the exposed animals become more aggressive, performing more aggressive behaviors towards the exposed animal and more redirected wall behaviors. • Physiology: • Our experiments indicate that perfusion of 20-HE over lateral antennule filaments causes large-scale excitation of individual olfactory receptor neurons (ORNs). • ORNs respond to 20-HE in a dose-dependent manner. These ORNs are broadly tuned and respond to amino acids common in their diet. Other steroids produce significantly lower levels of excitation and are not dose-dependent. • Biochemistry: • Sufficient total protein could be extracted from the following tissues: walking legs (WL), eyestalks (EY), intact antennules (IA), shaved antennule stalks (ST), aesthetascs (IA) and guard setae (GS). • SDS-PAGE gels indicate that the extracted protein has a MW that approximates that of the drosophila ecdysone receptor. • Western spot blots demonstrate that the following tissues (walking legs, eyestalk, intact antennules and antennule stalks) label with the antibody against the drosophila ecdysone receptor (EcR). • Biochemical data suggest that intact antennules and antennule shaved stalks have the EcR–further experiments will determine whether the membrane of aesthetascs and guard setae express the ecdysone receptor. • Our studies indicate that 20-HE is able to be perceived, affects social interactions, modulates internal aggressive states and neuromuscular transmission in Homarus americanus. This suggests that 20-HE acts both as a lobster hormone and a pheromone. REFERENCES Atema J, Cobb JS (1980) Social Behavior. In: The biology and management of lobsters. Vol 1. Cobb JS, Phillips BF (eds), pp 409-450. New York: Academic Press. Bolingbroke MB, Kass-Simon G (2001) 20-Hydroxyecdysone causes increased aggressiveness in female American lobsters, Homarus americanus. Horm and Behav 39:144-156. Breithaupt,T, Atema, J (1993) Evidence for the use of urine signals in agonistic encounters of the American lobster. Bio. Bull. 185:318. Breithaupt,T, Lindstrom, DP, Atema, J (1999). Urine release in freely moving catheterised lobsters (Homarus americanus) with reference to feeding and social activities. J exp Biol 202:837-844. Cromarty SI, Cobb JS, Kass-Simon G (1991) Behavioral analysis of the escape response in the juvenile lobster Homarus americanus over the molt cycle. J exp Biol 158:565-581. Cromarty SI and Kass-Simon G (1998) Differential effects of a molting hormone, 20-Hydroxyecdysone, on the neuromuscular junctions of the claw opener and abdominal flexor muscles of the American lobster. Comp Biochem Physiol 120(2):289-300. Derby CD (1995) Single unit electrophysiological recording techniques from crustacean chemoreceptor neurons. In: Spielman AI, Brand JG (eds) CRC Handbook on experimental cell biology of taste and olfaction: Current techniques and protocols. CRC, Boca Raton, FL, pp 241-250. Schwanke ML, Cobb JS, Kass-Simon G (1983) Physiological evidence for a blood-borne factor affecting aggressive behaviors in lobsters. Am Zool 23:947A. Schwanke ML, Cobb JS, Kass-Simon G (1990) Synaptic plasticity and humoral modulation of neuromuscular transmission in the lobster claw opener during the molt cycle. Comp Biochem Physiol 97C:143-149. Scrivener JCE (1971) Agonistic behavior of the American lobster, Homarus americanus (Milne-Edwards) Fish Res. Bd Can Tech Rep 235:1-128. Snyder MJ, Chang ES (1991a) Ecdysteroids in relation to the molt cycle of the American lobster, Homarus americanus. I. Hemolymph Titers and Metabolites. Gen Comp Endocrin 81:133-145. Snyder MJ, Chang ES (1991b) Ecdysteroids in relation to the molt cycle of the American lobster, Homarus americanus. II. Excretion of Metabolites. Gen Comp Endocrin 83:118-131. Talbot WS, Swyryd, EA, Hogness, DS (1993). Drosophila tissues with different metamorphic responses to ecdysone express different receptor isoforms. Cell 73:1323-1337. Tamm GR, Cobb JS (1978) Behavior and the crustacean molt cycle: Changes in aggression of Homarus americanus. Science 200:79-81. • PHYSIOLOGY METHODS • The lateral antennular filament was excised and inserted into a Teflon tube attached to an olfactometer in which artificial seawater (ASW) was flowing at 10 cm/s. • The proximal end of the antennule was inserted into a separate recording chamber containing Homarus saline. Several annuli were removed from the cut proximal end of the filament, thus exposing both the antennular nerve and artery. • The artery was then cannulated, and the preparation was perfused with oxygenated Homarus saline delivered directly into the artery through a glass cannula at a flow rate of 0.4–0.8 ml/min (see Derby, 1995). • Chemical stimuli were injected into the ASW flowing over the aesthetasc sensilla by activation of an electronic valve. The electronic valve allowed for a determination of the beginning of the stimulation period and response latency. Each odorant trial was followed by an ASW injection wash followed by a 2 minute inter-trial period. • Single-unit responses were recorded extracellularly from the exposed axons of ORNs in the saline dish by using fine-tipped glass suction electrodes (tip inner diameter ~ 2–5 µm), coupled to a differential AC amplifier. • Activity was monitored on-line and recorded on a microcomputer in digitized form using a Powerlab® interface and 5.4.2 software (AD Instruments). Spikes were sorted according to their waveforms using Spike Histogram Software of the Chart 5.4.2 package (AD Instruments). • For each stimulus and neuron, the numbers of spikes during 2000-ms periods following initiation of the response were counted as measures of the response intensity. • Dose dependence was evaluated with regression analysis (Excel). • “Best cells” are defined as those ORNs that have a maximum response to a particular odorant. Mixture search odorants are dose-dependent ACKNOWLEDGEMENTS The monoclonal antibody DDA2.7 was developed by Carl Thummel and was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by the University of Iowa, Department of Biological Sciences, Iowa City, IA 52242. Grant# IBN0234569 (GKS) Grant# IBN0234555 (SC)






