Download
1 / 44
440 likes | 575 Views
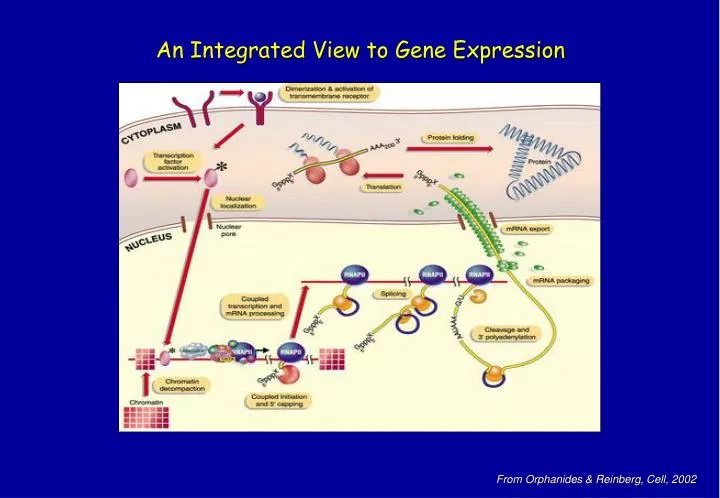
An Integrated View to Gene Expression. From Orphanides & Reinberg, Cell, 2002. Biochemistry of Transcription and Chromatin. In essence, gene transcription is a simply process, that can be divided into individual steps. Margaritis, Holstege, Cell 2008. “All information to correctly express a

E N D
An Integrated View to Gene Expression From Orphanides & Reinberg, Cell, 2002
In essence, gene transcription is a simply process, that can be divided into individual steps Margaritis, Holstege, Cell 2008
. “All information to correctly express a protein-encoding gene is in its sequence” promoter locus control region boundary element boundary element -500 -40 -4000 Gene X matrix attachment region matrix attachment region enhancers distal promoter core promoter enhancer
What is the “gene expression” parts list for a simple eukaryote? A simple eukaryote: the yeast Saccharomyces cerevisiae 6200 genes (5% are spliced), non-coding transcripts but no microRNAs ± 250 gene-specific transcription factors ± 150 chromatin remodeling and modifying factors ~65 general transcription machinery proteins ~20 transcription elongation proteins ± 800 “other” factors involved in regulation: such as 117 protein kinases, ±200 components of the ubiquitin machinery, mRNA splicing proteins, mRNA export factors
What is the “gene expression” parts list for a complex eukaryote? A more complex eukaryote, Homo sapiens: 22,000 mRNA genes (95% are spliced), ~1,000 microRNAs, 5,000-10,000 long non-coding RNAs ~2,500 gene-specific transcription factors ~1,000 chromatin proteins ~75 general transcription machinery proteins ~50 transcription elongation proteins “other” factors involved in regulation: such as ~500 protein kinases, ±1200 components of the ubiquitin machinery, mRNA splicing proteins, mRNA export factors
Gene-specific transcription factors require “Cofactors” (activators or repressors) to mediate their regulatory effects “Cofactors” Mediator Chromatin modifiers, e.g. HATs, HDACs, PRMTs, KMTs, KDMs Chromatin remodellers, e.g. ATP-dependent enzymes, nuclear lamina Histone binders, e.g. histone chaperones, HP1, elongation factors 19S proteasomal particle “upstream activator” “basal machinery” p50 p65 NF-kB TATA Interleukin 2 -40 +40 anywhere promoter-distal core promoter gene-specific general or basal
. “All information to correctly express a protein-encoding gene is in its sequence” promoter locus control region boundary element boundary element -500 -40 -4000 Gene X matrix attachment region matrix attachment region enhancers distal promoter core promoter enhancer but ....... it is very difficult to find functional elements by DNA sequence gazing
Promoter (~50 bp) recognition How to locate a core promoter? Accessibility The example of yeast 6,200 core promoters= 310,000 bp 310,000 in 12,400,000 bp 1 to 40 windows of 50 bp The example of human chromosome 22 400 core promoters= 400 x 50 bp= 2,000 bp 2,000 in 48,000,000 bp 1 to 24,000 windows of 50 bp
Location of core promoter by genomic sequencing of mRNA ends Sharp or focused pol II promoters: 10-20% of total contain a TATA-box Broad type of pol II promoters: 50-70% of total reside in a CpG islands Sandelin et al 2007 Nature Reviews Genet. 8, 424
General classification ofpol II promoters Sandelin et al Nat Rev Genet 8 (2007); Tora and Timmers TiBS 35 (2010)
TFIID binds both modified chromatin and promoter DNA DNA contacts: TBP TATA TAF1/2 INR TAF6/9 DPE Protein contacts: TAF1 H3-K9,14ac; H4-Kac TAF3 H3-K4me3 Jacobson et al. Science 288 (2000) Vermeulen et al. Cell 131 (2007) Guenther et al. Cell 130 (2007)
MTE BRE Five distinct DNA sequences in focused core promoters can attract the basal machinery “basal machinery” TATA INR DPE -38/-32 -30/-24 -2/+5 +18/+27 +28/+32 SSRCGCC TATAWAAR YYANWYY CSARCSS RGWYV ... but they are functionally redundant and are present in a small set of promoters.....
“Back to the seventies” • Paradigm for mRNA synthesis: bacterial enzyme (4 subunits) plus sigma-factor • Three nuclear RNA polymerases in human cells • Recombinant DNA cloning technology • Characterization of DNA viruses for animal cells (SV40, polyoma, adenovirus) Identification of the basal pol II machinery
Identification of the basal (or general) transcription factors by complementation in an in vitro pol II transcription assay using the TATA promoter of the adenovirus ML gene HeLa WCE phosphocellulose 0.6/0.5 1.0/0.85 0.04/0.1 0.35/0.3 [A] [B] [D] [C] TFIIB TFIIE TFIIF TFIIH TFIIA (B-TFIID) TFIID “all basal factors and pol II subunits are encoded by highly-conserved and unique genes” 33 basal factor polypeptides 13 RNA pol II peptides (+) 46 polypeptides
B Basal transcription factors assemble in a stepwise fashion -40 -30 -20 -10 +1 +10 pol II 34 74 F 56 E 30 TBP TATA H Promoter recognition Promoter opening Abortive Initiation Productive Initiation TBP-TFIID TFIIB TFIIF TFIIE TFIIH TFIIH (TFIIH)
MTE BRE Core promoter elements are bound by basal transcription factors “basal machinery” TATA INR DPE -38/-32 -30/-24 -2/+5 +18/+27 +28/+32 SSRCGCC TATAWAAR YYANWYY CSARCSS RGWYV TFIID TFIIB
. Properties of TBP and TFIID • TBP/TFIID: • 38-kDa in humans; 27-kDa in yeast • TATA-binding protein (TBP) is the major sequence-specific DNA binding component of TFIID • TBP is (always) associated in large multi-protein complexes • two complexes, TFIID and B-TFIID, support pol II transcription • 13 TBP-associated factors (TAFs) are in TFIID and B-TFIID has only one TAF • DNA binding by TBP (and TFIID) is slow and has limited sequence selectivity • TBP consists of a divergent N-terminus and a highly conserved C-terminal domain of 180 aa • (almost) all functions reside in this conserved domain • TBP is sufficient for basal transcription in vitro • activated transcription in vitro requires TAFs as cofactors • TAFs have been found to interact with many transcription factors • TAFs are not essential for transcription activation in vivo
TBP TBP TBP TBP BDP TBP distribution in human cell lines Nucleoplasm BTAF1/ Mot1p TAFI-48 TAFI-110 B-TFIID SL-1 TAFI-63 TAFI-41 Nucleolus pol II pol I TBP pol II pol III TFIID TFIIIB BRF1 TBP TBP 13 TAFs Peri-nucleolar
TBP-TAF complexes • TFIID 13 TAFs (250-18 kDa) plusTBP pol II • B-TFIID BTAF1(Mot1p) plusTBP pol II • SL 1 4 TAFs (110, 63, 48, 41) plusTBP pol I • TFIIIB 2 TAFs (150, 90 kDa) plusTBP pol III
TATA GeneX “basal machinery” Proteins directly affecting basal transcription by pol II TBP-associated factors: TAF1 can inhibit TATA-binding recognizes INR-element HAT activity ? double bromo-domain binding acetylated nucleosomes TAF2 recognizes INR-element TAF3 recognizes trimethylated lysine-4 of histone H3 involved in muscle differentiation (?) TAF4b selective for TGF signaling TAF6/9 contacts DPE BTAF1/MOT1p dissociates TBP-NC2-TATA complex dependent on ATP relaxed DNA binding specificity NC2 blocks TFIIA/TFIIB entry to preinitiation complex, induces TBP sliding on DNA relaxed DNA binding specificity TFIIA prevents action of repressors like NC2 increased stability of DNA interaction
. • RNA polymerase II: • consists of 12 subunits; all but one (RPB4) are essential • RBP1 and RBP2 are homologous to b’ and b of RNAP • some subunits (RBP5, -6, -10a, -10b) shared with pol I and pol III RPB1 ’ RBP2 RBP3 ”” RBP4 RBP5 RBP6 RBP7 RBP8 RBP9 RBP10a RBP10b RBP11 Nobel Prize Chemistry 2006: Kornberg "for his studies of the molecular basis of eukaryotic transcription" Speaker CHAINS 28/11/2011, Maarssen
. • RNA polymerase II: • has little intrinsic proofreading activity • low elongation efficiency, but high processivity • promoter-specific initiation requires ATP hydrolysis • RBP1 subunit contains unique heptad repeat structure (YSPTSPS) forming the so-called CTD (52 repeats in mammals) • CTD is target for kinases and phosphatases • CTD of elongating pol II is hyper phosphorylated • transcription cycle involves CTD-phosphorylation cycle • truncation of CTD by 50% gives conditional phenotypes in yeast • CTD is involved in many protein interactions (mRNA processing factors, coactivators)
. Cramer, Bushnell and Kornberg (2001) Science 292, 1863
. Gnatt et al. (2001) Science 292, 1876
. • RNA polymerase II: • has little intrinsic proofreading activity • low elongation efficiency, but high processivity • promoter-specific initiation requires ATP hydrolysis • CTD is target for kinases and phosphatases • CTD of elongating pol II is hyper phosphorylated • transcription cycle involves CTD-phosphorylation cycle • truncation of CTD by 50% gives conditional phenotypes in yeast • CTD is involved in many protein interactions (mRNA processing factors, coactivators)
Ser5-P Ser7-P Ser2-P The CTD is differentially phosphorylated during transcription consensus CTD-repeat: YSPTSPS CTD-kinases: cdk7/cyclinH of TFIIH (Ser5/7) cdk8/cyclinC of Mediator (Ser5) cdk9/cyclinT of PTEFb (Ser2) CTK1 (Ser2) CTD-phosphatases: FCP1 (Ser2) Ssu72 (Ser5)
. Discovery of cofactors • RNA polymerase II: • has little intrinsic proofreading activity • low elongation efficiency, but high processivity • promoter-specific initiation requires ATP hydrolysis • CTD is target for kinases and phosphatases • CTD of elongating pol II is hyper phosphorylated • transcription cycle involves CTD-phosphorylation cycle • truncation of CTD by 50% gives conditional phenotypes in yeast • CTD is involved in many protein interactions (mRNA processing factors, coactivators)
The phosphorylated CTD attracts mRNA processing factors mRNA processing: capping splicing poly(A)-tail addition
. Discovery of cofactors • RNA polymerase II: • has little intrinsic proofreading activity • low elongation efficiency, but high processivity • promoter-specific initiation requires ATP hydrolysis • CTD is target for kinases and phosphatases • CTD of elongating pol II is hyper phosphorylated • transcription cycle involves CTD-phosphorylation cycle • truncation of CTD by 50% gives conditional phenotypes in yeast • CTD is involved in many protein interactions (mRNA processing factors, coactivators)
SRB proteins were identified by genetic means in yeast the CTD of yeast pol II carries 26 heptad repeats deletion to less than 12 repeats gives rise to conditional phenotypes (inositol- auxotrophy, cold-sensitivity) yeast cells with less than 10 repeats are non-viable mutations in SRB (Suppressor of RNA polymerase B) genes restore viability biochemical purification showed that SRB proteins are associated in a multi- subunit complex termed the Mediator, which can bind directly to pol II
Mediator also known as ARC, SMMC, NAT, PC2, DRIP, TRAP complex
The Mediator complex Conserved from yeast to mammals Associated with RNA pol II Required for transcription nearly all protein-coding genes Required for response to activators
Mediator has at least two functions: required for the response to activators stimulates basal txpn levels Malik and Roeder (2005) Trends Biochem. Sci. 30, 256
Signal transduction? Negative components Enzymatic activities Opposite effects on genes upon deleting different components? Mediator mechanism
Transcription of HIV is controlled at the level of elongation
10-20% of genes contain paused pol II around 20-40 bp of TSS
Transcription elongation factors enhance the processivity of RNA polymerase II • TFIIS re-activates paused pol II by inducing exonucleolytic cleavage • Elongin (trimeric complex of elongin A , B and C) decreases Km • ELL • TFIIF • FACT (facilitates chromatin transcription) dissociates • nucleosomes upon transcription • DSIF (DRB sensitivity inducing factor) counteracts NELF • (negative elongation factor)-pausing of pol II • P-TEFb (cdk9/cyclinT kinase) phosphorylates the CTD on Ser2 • PAF1 complex • Spt6 helps transcription through chromatin by binding dissociated H3 • ....
Concepts in transcription regulation: transcription factor accessibility and recruitment are important regulatory mechanisms basal transcription factors are non-redundant and non-modular and they function position- and orientation-dependent gene-specific factors are (often) redundant and dimeric and they function relatively position- and orientation-independent gene-specific factors and cofactors are part of multi-gene families and they contain separate modules for DNA- or factor-binding and for transcription regulation chromatin-dependent cofactors are large protein complexes chromatin-dependent complexes can have both positive and negative effects on transcription synergism, protein-modification and abundance are important regulatory mechanisms for gene-specific factors .