Download
1 / 1
10 likes | 156 Views
Déjà Vu Communities: When Invaders Meet Again Melanie Northrup, Christy Cincotta, Mark June-Wells, Jonathan Lansey, Kunj Patel, Apiwat Romayanantakit, Claus Holzapfel Rutgers University Newark, Dept. of Biological Sciences, Newark NJ 07102-1811 contact: holzapfe@andromeda.rutgers.edu.

E N D
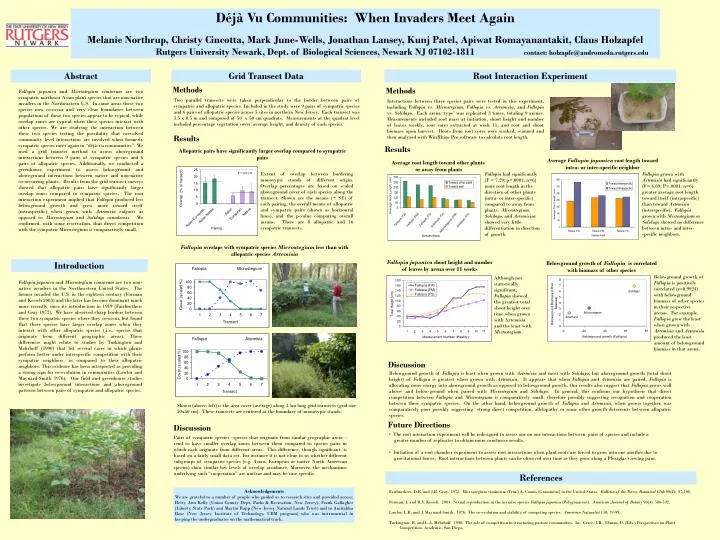
Déjà Vu Communities: When Invaders Meet Again Melanie Northrup, Christy Cincotta, Mark June-Wells, Jonathan Lansey, Kunj Patel, Apiwat Romayanantakit, Claus Holzapfel Rutgers University Newark, Dept. of Biological Sciences, Newark NJ 07102-1811 contact: holzapfe@andromeda.rutgers.edu Abstract Grid Transect Data Root Interaction Experiment Methods Methods Fallopia japonica and Microstegium vimineum are two sympatric northeast Asian plant species that are non-native invaders in the Northeastern U.S. In some areas these two species now co-occur and very clear boundaries between populations of these two species appear to be typical, while overlap zones are typical when these species interact with other species. We are studying the interaction between these two species testing the possibility that coevolved community level interactions are involved when formerly sympatric species meet again in “déjà-vu communities”. We used a grid transect method to assessaboveground interactions between 9 pairs of sympatric species and 6 pairs of allopatric species. Additionally, we conducted a greenhouse experiment to assess belowground and aboveground interactions between native and non-native co-occurring plants. Results from the grid transect surveys showed that allopatric pairs have significantly larger overlap zones compared to sympatric species. The root interaction experiment implied that Fallopia producedless belowground growth and grew more toward itself (intraspecific) when grown with Artemisia vulgaris as opposed to Microstegium and Solidago canadensis. We confirmed, with some reservation, that direct competition with the sympatric Microstegium is comparatively small. Two parallel transects were taken perpendicular to the border between pairs of sympatric and allopatric species. Included in the study were 9 pairs of sympatric species and 6 pairs of allopatric species across 5 sites in northern New Jersey. Each transect was 3.5 x 0.5 m and composed of 50 x 50 cm quadrats. Measurements at the quadrat level included percentage vegetation cover, average height, and density of each species. Interactions between three species pairs were tested in this experiment, including Fallopia vs. Microstegium,Fallopia vs. Artemisia, and Fallopia vs. Solidago. Each arena ‘type’ was replicated 3 times, totaling 9 arenas. Measurements included root mass at initiation, shoot height and number of leaves weekly, root cores extracted at week 11, and root and shoot biomass upon harvest. Roots from root cores were washed, scanned and then analyzed with WinRhizo Pro software to calculate root length. Results Results Allopatric pairs have significantly larger overlap compared to sympatric pairs Average Fallopia japonica root length toward intra- or inter-specific neighbor Average root length toward other plants or away from plants P=0.0130 Fallopia had significantly (F = 7.29; p<.0001; n=6) more root length in the direction of other plants (intra- or inter-specific) compared to away from plants. Microstegium, Solidago, and Artemisisa showed very little differentiation in direction of growth. Fallopia grown with Artemisia had significantly (F= 4.69; P<.0001; n=6) greater average root length toward itself (intraspecific) than toward Artemisia (interspecific). Fallopia grown with Microstegium or Solidago showed no difference between intra- and inter-specific neighbors. Fallopia overlaps with sympatric species Microstegium less than with allopatric species Artemisia Fallopia japonica shoot height and number of leaves by arena over 11 weeks Introduction Belowground growth of Fallopia is correlated with biomass of other species Belowground growth of Fallopia is positively correlated (r=0.9924) with belowground biomass of other species in their respective arenas. For example, Fallopia grew the least when grown with Artemisia and Artemisia produced the least amount of belowground biomass in that arena. Although not statistically significant, Fallopia showed the greatest total shoot height over time when grown with Artemisia and the least with Microstegium. Fallopia japonica and Microstegium vimineum are two non-native invaders in the Northeastern United States. The former invaded the U.S. in the eighteen century (Forman and Kesseli 2003) and the later has become dominant much more recently since its introduction in 1919 (Fairbrothers and Gray 1972). We have observed sharp borders between these two sympatric species where they co-occur, but found that these species have larger overlap zones when they interact with other allopatric species (.i.e., species that originate from different geographic areas). These differences might relate to studies by Turkington and Mehrhoff (1990) that list several cases in which plants perform better under interspecific competition with their sympatric neighbors, as compared to their allopatric neighbors. This evidence has been interpreted as providing a strong sign for co-evolution in communities (Lawlor and Maynard-Smith 1976). Our field and greenhouse studies investigate belowground interactions and aboveground patterns between pairs of sympatric and allopatric species. Discussion Belowground growth of Fallopia is least when grown with Artemisia and most with Solidago, but aboveground growth (total shoot height) of Fallopia is greatest when grown with Artemisia. It appears that when Fallopia and Artemisia are paired, Fallopia is allocating more energy into aboveground growth as opposed to belowground growth. Our results also suggest that Fallopia grows well above- and below-ground when paired with Microstegium and Solidago. In general, this confirms our hypothesis that direct competition between Fallopia and Microstegium is comparatively small, therefore possibly suggesting recognition and cooperation between these sympatric species. On the other hand, belowground growth of Fallopia and Artemisia, when grown together, was comparatively poor possibly suggesting strong direct competition, allelopathy or some other growth deterrents between allopatric species. Shown (above, left) is the area cover (average) along 3.5m long grid transects (grid size 50x50 cm). These transects are centered at the boundary of monotypic stands. Future Directions Discussion • The root interaction experiment will be redesigned to assess one on one interactions between pairs of species and include a • greater number of replicates to obtain more conclusive results. • Initiation of a root chamber experiment to assess root interactions when plant roots are forced to grow into one another due to • gravitational forces. Root interactions between plants can be observed over time as they grow along a Plexiglas viewing pane. Pairs of sympatric species –species that originate from similar geographic areas – tend to have smaller overlap zones between them compared to species pairs in which each originate from different areas. This difference, though significant, is based on a fairly small data set. For instance it is not clear to us whether different subgroups of sympatric species (e.g. Asian, European or native North American species) show similar low levels of overlap avoidance. Moreover, the mechanisms underlying such “cooperation” are unclear and may be case specific. References Acknowledgements We are grateful to a number of people who guided us to research sites and provided access: Betty Ann Kelly (Union County Dept. Parks & Recreation, New Jersey), Frank Gallagher (Liberty State Park) and Martin Rapp (New Jersey Natural Lands Trust) and to Amitabha Bose (New Jersey Institute of Technology, UBM program) who was instrumental in keeping the undergraduates on the mathematical track. Fairbrothers, D.E. and J.R. Gray. 1972. Microstegium vimineum (Trin.) A. Camus (Gramineae) in the United States. Bulletin of the Torrey Botanical Club 99(2): 97-100. Forman, J. and R.V. Kesseli. 2003. Sexual reproduction in the invasive species Fallopia japonica (Polygonaceae). American Journal of Botany 90(4): 586-592. Lawlor, L.R. and J. Maynard-Smith. 1976. The co-evolution and stability of competing species. American Naturalist 110: 79-99. Turkington, R. and L.A. Mehrhoff. 1990. The role of competition in structuring pasture communities. In: Grace, J.B., Tilman, D. (Eds.) Perspectives on Plant Competition, Academic, San Diego.