Download

1 / 201
2.09k likes | 2.81k Views
Chapter12 The genetic Code and Transcription. Genetic information transferred to RNA by the process of transcription. Electron micrograph visualizing the process of transcription. 12.1 Characteristics of the genetic code 12.2 The triplet Nature of the code

E N D
Chapter12 The genetic Code and Transcription Genetic information transferred to RNA by the process of transcription. Electron micrograph visualizing the process of transcription.
12.1 Characteristics of the genetic code 12.2 The triplet Nature of the code 12.3 Deciphering the code and 12.4 Coding dictionary 12.5 Confirming the code using phage MS2 12.6 The universality of the genetic code 12.7 Transcription : DNA-dependent RNA synthesis 12.8 RNA polymerase 12.9Transcription in eukaryotes 12.10 Intervening sequences and split genes 12.11 RNA editing
Information in the linear sequence of deoxyribonucleotides Medium transcribed from DNA Ended products
12.1 Characteristics of the genetic code 1. The genetic code is written in linear form, using the ribonucleotide bases that compose mRNA molecules as"letters." The ribonucleotide sequence is derived from the complementary nucleotide bases in DNA, 2. Each "word" within the mRNA contains three ribonucleotide letter. Each group of three ribonucleotides, called a codon, specifies one amino acid; the code is thus a triplet. 3. The code is unambiguous, meaning that each triplet specifies only a single amino acid.
4. The code is degenerate; that is, a given amino acid can be specified by more than one triplet codon. This is the case for l8 of the 20 amino acids. 5. The code contains "start" and "stop" signals. Certain triplets that initiate and terminate translation. 6. No internal punctuation ("commas") is used in the code. Thus, the code is said to be commaless. Once translation of mRNA begins, the codons are read one after the other with no breaks between them. 7, The code is nonoverlapping. Once translation commences, any single ribonucleotide at a specific location within the mRNA is part of only one triplet, 8. The code is nearly universal. With only minor exceptions. a single coding dictionary is used by almost all viruses, prokaryotes, archaea, and eukaryotes.
what do we know ? • DNA transcription RNA translation protein • relationships are colinear • nonoverlapping code (1 mutation, 1 amino acid)
12.2 The triplet Nature of the code In the early 1960s. Sidney Brenner argued on theoretical grounds that the code must be a triplet since three-letter words represent the minimal use of four 1etters to specify 20 amino acids. • how many letters in DNA code / amino acid “words”? • 4 bases: A, T (U), C, G • 20 amino acids • if... • 1 base / aa 4 aa possible (41) • 2 bases / aa 16 aa possible (42) • 3 bases / aa 64 aa possible (43)... minimum # • 4 bases / aa 256 aa possible (44)
Using phage T4 by Francis Crick and his co1leagues who studied frameshift mutation experimental evidence supporting the triplet nature of the code • The addition or deletion of one or more nucleotides The gain or the loss of one or more letters shifts the frame of reading during translation. • But when three nucleotides were involved. the frame of reading was reestablished (Figure l2--2).
12.3 Deciphering the code • In l96l, Marshall Nirenberg and J. Heinrich Matthaei characterized the first specific coding sequences. which served as a cornerstone for the complete analysis of the genetic code • dependent on the use of two experimental tools, an in vitro (cell-free) protein-synthesizing system, and an enzyme, polynucleotide p phosphorylase 多核苷酸磷酸化酶, 多核苷酸转核苷酰酶, which allowed the production of synthetic mRNAs
Cell-Free Polypeptide Synthesis • 1955, bacterial Enzyme • 1961, mRNA had yet to be isolated • made synthetic mRNA=nucleotides + polynucleotide phosphorylase enzyme • nucleotide incorporation random single strand • poly(U) mRNA + E. coli machinery in vitro only phenylalanine (Phe) residues
the cell-free system (in vitro)for protein synthesis and the availability of synthetic mRNAs provided a means Of deciphering the ribonucleotide composition of various triplets encoding specific amino acids.
In contrast to RNA polymerase, polynucleotide phosphorylase requires no DNA template. • As a result, each addition of a ribonucleotide is random, based on the relative concentration of the four ribonucleoside diphosphates added to the reaction mixtures. The probability of the insertion of a specific ribonucleotide is proportional to the availability of that molecule, relative to other available ri-bonucleotides. • This point is absolutely critical to understanding the work of Nirenberg and others in the ensuing discussion.
The Use of Homopolymers • In their initial experiments. Nirenberg and Matthaei synthesized RNA homoplymers, each consisting of only one type of ribonucleotjde. • UUUUUU. AAAAAA... CCCCCC.. Or GGGGGG... In testing each mRNA, they were able to determine which. if any amino acids were incorporated into newly synthesized proteins,with labeled one of the 20 amino acids added to the in vitro system. Since three Us, As,and Cs, only one possible sequence, the actual codon was identified. • UUU---- Phe CCC---Pro • AAA----Lys GGG---?gly
Mixed Copolymers • With these techniques in hand. Nirenberg and Matthaei, and Ochoa and coworkers turned to the use of RNA heteropolymers. In this technique. two or more different ribonucleoside diphosphates are added in combination to form the artificial message. • The researchers reasoned that if they knew the relative proportion of each type of ribonucleoside diphosphate, they could predict the frequency of any particular triplet codon occurring in the synthetic mRNA. If they then added the mRNA to the cell-free system and ascertained the percentage of any particular amino acid present in the new protein, they could analyze the results and predict the composition of triplets specifying specific amino acids.shown in Fig12-4.
Suppose A:C in a ratio of 1:5 11.6+2.3 57.9+11.6
GENETIC CODE • mixed nucleotides in known ratios... e.g. ¾ U + ¼ G...
GENETIC CODE • some outcomes not unique, can’t determine code
GENETIC CODE • ¾ U + ¼ G 6 amino acids...
GENETIC CODE AA Phe Leu Cys Val Trp Gly Val Gly Obs. 1.00 0.37 0.35 0.36 0.14 0.12
The Triplet Binding Assay • It was not long before more advanced techniques were developed. In I 964. Nirenberg and Philip Leder developed the triple binding assay, which led to specific assignments of triplets.The technique took advantage of the observation that ribosomes, when presented with an RNA sequence as short as three ribonuchotides, will bind to it and form a complex similar to that found in vivo. The triplet acts like a codon in mRNA, attracting the complementary sequence within tRNA (Figure l2-5). Such a triplet sequence in tRNA that is complementary to a codon of mRNA is known as an anticodon.
Although it was not yet feasible to chemically synthesize long stretches of RNA. triplets of known sequence could be synthesized in the 1aboratory to serve as templates. All that was needed was a method to determine which tRNA-amino acid was bound to the triplet RNA--ribosome complex.
The test system devised was quite simple. The amino acid to be tested was made radioactive, and a charged tRNA was produced. Because code compositions were known, researchers could narrow the decision as to which amino acids should be tested for each specific triplet.
The radioactively charged tRNA, the RNA triplet, and ribosomes are incubated together on a nitrocellulose filter, which retains the larger ribosomes but not the other smaller components, such as charged tRNA. If radioactivity is not retained on the filter, an incorrect amino acid has been tested. If radioactivity remains on the filter, it is retained because the charged tRNA has bound to the triplet associated with the ribosome. In such a case, a specific codon assignment can be made.
Table 12.2 Amino Acids assignment to specific trinucleotides derived from the triplet binding assay Trinucleotides Amino Acids UGU UGC Cysteine GAA GAG Glutamic acid AUU AUA AUG Isoleucine UUA UUG CUU CUC CUA CUG Leucine AAA AAG Lysine AUG methionine UUU UUC Phenylalanine CCU CCC CCG CCA Proline UCU UCC UCA UCG Serine 26 triplet for 9 amino acids
Table 12.2, for example, shows 26 triplets assigned to 9 amino acids. However, in some cases, the degree of triplet binding was inefficient, and assignments were not possible. Eventually, about 50 of the 64 triplets were assigned. These specific assignments of triplets to amino acids led to two major conelusions. First, the code degenerate; that is, one amino acid can be specified by more than one triplet. Second, the code is unambiguous; that is, a single triplet specifies only one amino acid.
Repeating Copolymers • Yet another innovative technique used to decipher the genetic code was developed in the early l960s by Gobind Khorana, who chemically synthesized long RNA molecules consisting of short sequences repeated many times. First, he created shorter sequences (eg.. di-. tri-. and tetranucleotides), which were then replicated many times and finally joined enzymatically to form the long polynucleotides. As shown in Figure l2-6. a dinucleotide made in this way is converted to a mRNA with two repeating triplets. A trinucleotide is converted to a mRNA with three potential triplets, depending on the point at which initiation occurs, and a tetranuc1eotide creates four repeating triplets.
When these synthetic messages were added to a cell-free system. the predicted number of amino acids incorporated was upheld. Table 12.3. Repeating codon amino acids in Copolymer produced polypeptides UG UGU Cysteine GUG Valine AC ACA Threonine CAC Histine UUC UUC Phenylalianine UCU Serine CUU Leucine AUC AUC Isoleucine UCA Serine CAU Histine UACU UAU Tyrosine CUA Leucine UCU Serine AUC isoleucine
12.4 Coding dictionary • 3 stop codons • UAG = amber琥珀 • UAA = ochre赭石 • UGA = opal 乳白
genetic code is degenerate... >1 mRNA codon / aa • 1 Trp • 6 Ser • 2 ways... • >1 tRNA • >1 codon wobble...
wobble at 5' site of anticodon • 1 tRNA anticodon >1 mRNA codon
wobble rules • 1 tRNA anticodon >1 mRNA codon
3 different Ser-tRNASer (+1 more)... isoaccepting tRNA • + wobble (x2) = 6 possible codons
12.5 Confirming the code using phage MS2 • These individua1 principles have been confirmed by the detailed analysis of the RNA-containing bacteriophage MS2 by Waller Fiers and his co-workers • MS2- 3500nt, make up only 3 three genes Amino acid of the coat protein sequenced 1970 By 1976,other two genes
Fiers1972年对MS2 phage外壳蛋白基因及RNA分子进行了测序。图11-45为MS2一部分核苷酸序列及相对应的氨基酸。从图可以看出:
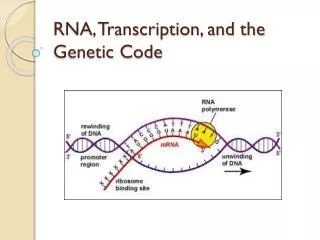
12.7 Transcription : DNA-dependent RNA synthesis • 现在将作为转录模板的DNA单链称为模板链或反义链(antisen strand), • 非模板链称为有义链(sense strand)或编码链。RNA Polymerase prokaryote eukaryotes • Promoter,template binding,with transcription factor (initiation) prokaryote Eukaryotes • Termination • Maturation
RNA Polymerase • RNA polymerases transcribe all types of RNA • in prokaryotes... • RNA pol _ all types of RNA • polycistronic mRNA
RNA Polymerase From E.coli shown to consist of subunits,designated as holoenzyme
The major form is designated as σ70, based on its molecular weight of 70 kilodaltons (kDa). While the promoters of most bacterial genes recognize this form, several alternative forms of RNA polymerase in E. coli have unique a subunits associated with them (e.g., σ28, σ32, σ38, and σ54). These recognize different promoter sequences and provide specificity to the initiation of transcription.
典型启动子的结构 -35 -10 转录起点 TTGACA 16-19bp TATAAT 5-9bp
Promoter,template binding, and the subunit • 一般长40-60bp,共有性序列(consensus sequences)。如图19-4所示,可分为识别(R,recognition)、结合(B, binding)和 起始(I, initiation)三个区段。 • 转录起始第一个碱基(通常标记位置为+1)最常见的是A; • 在-10bp附近有TATAAT一组共有序列,因为这段共有序列是Pribnow首先发现的,称为Pribnow盒(Pribnow box); • 在-35bp处又有TTGACA一组共有序列 。
-35序列又称为Sextama盒(Sextama box), • 其保守序列为(T82T84G78A65C54A45) • 其功能是: (1) 为RNA pol的识别位点。 σ亚基识别-35序列,为转录选择模板 (2)-35和-10序列的距离是稳定的,此与RNA pol的结构有关。
TRANSCRIPTION • initiation • 2 regions of homology among promoter sequences • consensus sequences = RNA pol binding sites