Download

1 / 3
30 likes | 56 Views
Otra estructura considerable estu00e1 formada por los llamados canales iu00f3nicos, que estu00e1n formados por proteu00ednas de canal y aceptan cierto trueque de masa. especialmente para el trueque con el agua para sobrepasar la barrera hidrofu00f3bica entre las dos capas de lu00edpidos de la membrana celular, estu00e1n presentes los llamados canales de agua (acuaporinas), que trabajan de manera semejante a los canales iu00f3nicos.

E N D
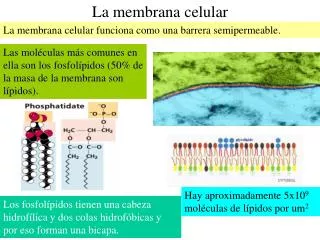
La membrana celular debe procesar muchas señales del ámbito y del interior de la célula para iniciar una respuesta molecular correcta. El acoplamiento de algunas sustancias mensajeras a la membrana, por ejemplo, puede desencadenar el desarrollo o la división de una célula. La membrana celular ha sido durante un largo tiempo el foco de la ciencia. por otro lado, en gran medida no estaba claro cómo se organizarían precisamente sus componentes individuales. Según un modelo obsoleto, las grasas y proteínas ancladas en la membrana debían desplazarse libremente y sin construcciones sólidas. Sólo en los años anteriores se ha demostrado la presencia de una organización en distintos espacios usando unas pocas proteínas. Además, la membrana celular es un espacio importante de comunicación. Las excitaciones eléctricas de las células se alargan a través de la membrana celular, lo cual es sustancial para la conducción de las células nerviosas y sus extensiones. Las funcionalidades de las células están controladas por señales extracelulares, la mayor parte de las cuales se reciben en los receptores de la membrana celular. la mayor parte de las proteínas ligadas a la membrana trabajan como receptores (sitios de unión) para algunos sustratos o ligandos y por lo general son muy específicas. en este texto describimos una metodología para evaluar la dinámica de reparación de la membrana celular en fibroblastos lesionados con láser de dos fotones, aunque esta metodología puede aplicarse a diferentes tipos de células para cuantificar la aptitud de resellado in vitro de la membrana plasmática. En este procedimiento, las células se incuban con FM4-64, un tinte lipófilo, impermeable a las células, que presenta una rápida fluorescencia al sumarse a los fosfolípidos cargados negativamente en el citoplasma al ingresar en la célula por medio de la lesión de la membrana (Figuras 2 y 3). La cuantificación de la fluorescencia del colorante adyacente a la lesión de la membrana permite monitorizar el tiempo que tarda la membrana celular en cerrarse. Para ilustrar la utilidad de este procedimiento, usamos fibroblastos de pacientes con disferlinopatía transfectados con plasma de disferlina de longitud total conjugado (DYSF) para evaluar el rescate de la reparación de la membrana celular. durante un largo tiempo, los estudiosos reflexionaron que los receptores nadan de forma individual y aleatoria en la capa lipídica de la membrana celular. no obstante, en un estudio publicado en 2010, Reth dió a conocer que los receptores de antígenos forman oligómeros: Se unen entre sí a una distancia de 10 a 20 nanómetros y forman aproximadamente 100 islas de receptores nanométricos en la área de la célula. La membrana celular está formada por una doble cubierta de lípidos y sólo tiene un espesor de 6 a 10 nanómetros. Los grupos lipofílicos de ámbas capas de lípidos se enfrentan entre sí y forman de esta forma una barrera hidrofóbica que no puede ser superada por los líquidos acuosos. Los lípidos de la cubierta externa están medianamente glicolados, los sacáridos pueden haberse juntado y unido con los lípidos para formar glicolípidos. Toda la membrana estará unida, pero por medio de la interacción no covalente de la cola hidrofóbica, no obstante, la estructura es más bien fluida y no está rígidamente fijada en su lugar. En condiciones fisiológicas, las moléculas fosfolípidas de la membrana celular se encuentran en estado cristalino líquido. esto significa que las moléculas de lípidos están www.membranacelular.com libres para difundir una rápida difusión lateral a lo largo de la capa y para mostrar dónde están presentes. no obstante, el trueque de moléculas fosfolípidas entre los folíolos intracelulares y extracelulares de la bicapa es un desarrollo muy lento. Las balsas de lípidos y caveolae son ejemplos de microdominio enriquecido con colesterol en la membrana celular. Además, una parte del lípido está en contacto directo con las proteínas de la membrana integral, que está estrechamente unida a la superficie de la proteína, se denomina envoltura lipídica anular; se comporta como una sección del complejo proteico. Todas las células se dividen por membranas en espacios de oposición cerrados (compartimentación). La membrana celular está compuesta por una bicapa lipídica donde se integran varias proteínas y algunas moléculas de colesterol. por lo tanto, forman una pared protectora contra sustancias polares o solubles en agua, mientras que las sustancias lipofílicas o no polares pueden penetrar de forma sencilla en la membrana. Los lípidos de la membrana plasmática son móviles, por ejemplo, la fluidez de la membrana es ocasionada por la movilidad de los lípidos. Los fosfolípidos tienen una cabeza hidrofílica y una cola hidrofóbica. Los fosfolípidos
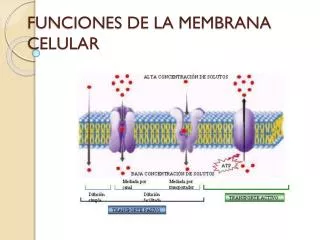
están con más grande continuidad en la membrana celular y forman la doble cubierta de la membrana, con los extremos de la cola sobresaliendo hacia el interior de esta cubierta. La membrana celular es semipermeable y por lo tanto permeable para algunas sustancias y no para otras. Al estar compuesto por una doble capa lipídica (componente principal: fosfolípido), es permeable a las moléculas lipofílicas (apolares). Las moléculas hidrofílicas o polares, de esta forma como los iones, los ácidos nucleicos, las proteínas y las partículas enormes sólo pueden pasar a través de la membrana a través de distintos mecanismos de transporte. también analizaron la función del canal iónico usando procedimientos electrofisiológicos. La proteína VRAC consiste en seis subunidades. Dispuestos alrededor de un eje habitual, estos forman el canal. La proteína, que se encuentra en la membrana celular, tiene dentro una pequeña zona que se proyecta hacia el exterior y una gran sector que está dentro de la célula. El carácter lipofílico de la membrana celular facilita separar dos ambientes acuosos y evadir la mezcla. Los iones, las moléculas fuertemente polares y las moléculas más importantes no tienen la posibilidad de o sólo tienen la posibilidad de pasar con dificultad a través de la membrana. de esta manera, las células son capaces de canjear selectivamente sustancias entre las dos millas a través de canales y transportadores integrados en la bicapa lipídica: Esto permite mantener las diferencias de concentración (nota: no debe usarse el término "gradiente") y, por ejemplo, amontonar un potencial de membrana. Los receptores dentro de la membrana permiten la transmisión de señales al entorno intracelular. estudiosos dirigidos por el Prof. Martin Kerschensteiner, Director del centro de Neuroinmunología Clínica de la LMU, y el instructor TUM Thomas Misgeld del centro de Biología Celular Neuronal han identificado un mecanismo en el modelo animal de la esclerosis múltiple que puede conducir a la muerte de axones. Los científicos reportaron que la afluencia de calcio por medio de diminutas grietas en la membrana celular es responsable de esto. La membrana celular, al estar expuesta al ámbito externo, es un sitio sustancial de comunicación célula-célula. Como tal, una extensa variedad de receptores de proteínas y proteínas de identificación, como los antígenos, están presentes en la superficie de la membrana. Las funcionalidades de las proteínas de la membrana además pueden incluir el contacto célula-célula, el reconocimiento de la área, el contacto citoesquelético, la señalización, la actividad enzimática o el transporte de sustancias por medio de la membrana. Las sustancias que no pueden difundirse libremente (véase más abajo) se acumulan a ambos lados de las membranas, donde han llegado por difusión, y tienen que ser transportadas a través de la barrera de la membrana con sistemas de transporte activos, por ejemplo, aminoácidos, proteínas, colesterol y otros. Los movimientos descritos para la membrana celular y el citoplasma se aplican por igual a las membranas y al plasma de los orgánulos. La membrana celular separa el interior de la célula del exterior o de otras células y protege el núcleo celular, los orgánulos, el citoplasma y otras partes de la célula. más allá de su semipermeabilidad, la membrana puede dividir el fluido acuoso dentro de la célula del fluido acuoso fuera de la célula, inclusive a diferentes presiones osmóticas. 3ª Endocitosis: La endocitosis es el proceso por el cual las células absorben las moléculas devorándolas. La membrana plasmática produce una pequeña deformación hacia adentro llamada invaginación donde la sustancia debe ser transportada, la invaginación de las proteínas causadas en el exterior de la membrana celular trabaja como receptores y agrupación en depresiones que ocasionalmente promueven la acumulación de más proteínas y lípidos en el lado citosólico de la membrana. La deformación entonces se exprime de la membrana en el interior de la célula, creando una vesícula que tiene dentro la sustancia absorbida. La endocitosis es una forma de internalizar partículas sólidas ("nutrición celular o fagocitosis"), pequeñas moléculas e iones ("beber de células o pinocitosis") y macromoléculas. La endocitosis necesita energía y, por lo tanto, es una forma de transporte activo.
La estructura fué mencionada numerosas veces por distintos autores como ectoplasto ("de Vries", 1885), piel plasmática (piel plasmática, pimienta, 1877, 1891), capa de piel (capa de piel, pimienta, 1886, utilizada con un significado diferente por Hofmeister, 1867), membrana plasmática (pimienta, 1900), membrana plasmática, membrana plasmática, membrana citoplasmática, envoltura celular y membrana celular. Algunos autores que no creían que hubiera un límite servible permeable en la superficie de la célula prefirieron utilizar el término plasmalema (acuñado por Mast, 1924) para el exterior de la célula. Si las proteínas de la membrana acoplada son defectuosas, el sistema inmunitario puede no clasificar las células como tejido autólogo sino como tejido raro e inducir los ataques correspondientes. El síndrome antifosfolípido de la patología autoinmune (APS) transporta a una variación de la estructura de las membranas celulares de los glóbulos rojos ("eritrocitos") porque el sistema inmunitario lleva a la destrucción de las proteínas de la membrana asociadas con los fosfolípidos. La endocitosis y la exocitosis sirven para transportar sustancias que carecen de otros sistemas de transporte (por ejemplo, colesterol, proteínas). Estas sustancias se transportan en vesículas. Estos recipientes se forman a partir de la membrana celular o de la membrana de los orgánulos y vuelven a fusionarse con ellos luego de la liberación de la sustancia. El gradiente de concentración de sodio en las membranas celulares es importante para los procesos eléctricos, para los transportes activos (symporte, antiporte), el cuidado del volumen celular mediante la estabilización de la presión osmótica.