Download
1 / 1
20 likes | 137 Views
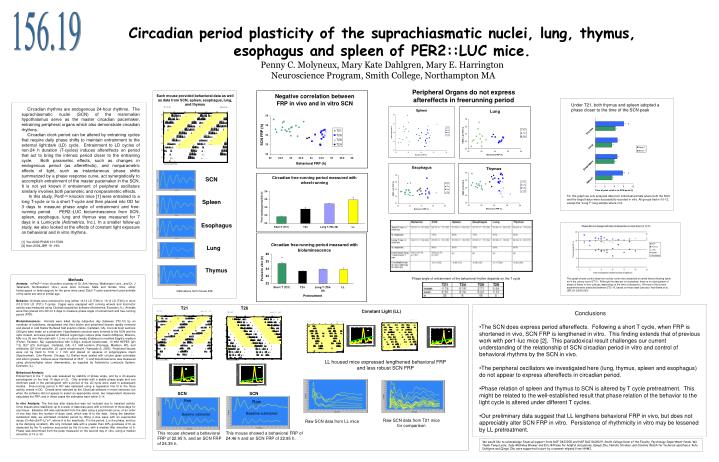
156.19. Circadian period plasticity of the suprachiasmatic nuclei, lung, thymus, esophagus and spleen of PER2::LUC mice. Penny C. Molyneux, Mary Kate Dahlgren, Mary E. Harrington Neuroscience Program, Smith College, Northampton MA. Peripheral Organs do not express

E N D
156.19 Circadian period plasticity of the suprachiasmatic nuclei, lung, thymus, esophagus and spleen of PER2::LUC mice.Penny C. Molyneux, Mary Kate Dahlgren, Mary E. Harrington Neuroscience Program, Smith College, Northampton MA Peripheral Organs do not express aftereffects in freerunning period Each mouse provided behavioral data as well as data from SCN, spleen, esophagus, lung, and thymus Under T21, both thymus and spleen adopted a phase closer to the time of the SCN peak Circadian rhythms are endogenous 24-hour rhythms. The suprachiasmatic nuclei (SCN) of the mammalian hypothalamus serve as the master circadian pacemaker, entraining peripheral organs which also demonstrate circadian rhythms. Circadian clock period can be altered by entraining cycles that require daily phase shifts to maintain entrainment to the external light:dark (LD) cycle. Entrainment to LD cycles of non-24 h duration (T-cycles) induces aftereffects on period that act to bring the intrinsic period closer to the entraining cycle. Both parametric effects, such as changes in endogenous period (as aftereffects), and nonparametric effects of light, such as instantaneous phase shifts summarized by a phase response curve, act synergistically to accomplish entrainment of the master pacemaker in the SCN. It is not yet known if entrainment of peripheral oscillators similarly involves both parametric and nonparametric effects. In this study, Per2Luc knockin mice [1] were entrained to a long T-cycle or to a short T-cycle and then placed into DD for 3 days to measure phase angle of entrainment and free-running period. PER2::LUC bioluminescence from SCN, spleen, esophagus, lung and thymus was measured for 7 days in a Lumicycle (Actimetrics, Inc.). In a smaller follow-up study, we also looked at the effects of constant light exposure on behavioral and in vitro rhythms. [1] Yoo 2000 PNAS 101:5339 [2] Aton 2004,JBR 19 :198; * * SCN For this graph we only analyzed data from individual animals where both the SCN and the target tissue were successfully recorded in vitro. All groups had n=10-12, except the “Long T” lung sample where n=4. Spleen Esophagus Lung * Thymus Phase angle of entrainment of the behavioral rhythm depends on the T cycle This graph shows control data from colony room mice dissected at varied times following lights on in the colony room (ZT 0). Although the data set is incomplete, there is no clear pattern of phase of these in vitro cultures depending on the time of dissection. All mice in the current experiment were dissected between ZT2-10, based on these data (see also Yoshikawa et al., JBR 20 (2005) 500). Methods Animals:mPer2Luc mice (founders courtesy of Dr. Erik Herzog, Washington Univ., and Dr. J Takahashi, Northwestern Univ.) were bred in-house. Male and female mice, either homozygous or heterozygous for the gene were used. Each T-cycle experiment used animals of the same sex and of similar age. Behavior: Animals were entrained to long (either 14:14 LD (T28) or 13:13 LD (T26)) or short (10.5:10.5 LD (T21)) T-cycles. Cages were equipped with running wheels and locomotor activity was measured using Clocklab acquisition software (Actimetrics, Evanston, IL). Animals were then placed into DD for 3 days to measure phase angle of entrainment and free-running period (FRP). Bioluminescence: Animals were killed during subjective day (between ZT2-10) by an overdose of halothane, decapitated and their brains and peripheral tissues rapidly removed and placed in cold Hanks’ Buffered Salt solution (Gibco, Carlsbad, CA). Coronal brain sections (300μm) were made on a vibratome. Hypothalamic sections were trimmed to the SCN and the optic chiasm, and were placed on Millicell organotypic culture plate inserts (Millipore, Billerica, MA) in a 35 mm Petri dish with 1.2 mL of culture media (Dulbecco’s modified Eagle’s medium (Fisher, Fairlawn, NJ) supplemented with 0.35g/L sodium bicarbonate, 10 mM HEPES (pH 7.2), B27 (2% Invitrogen, Carlsbad, CA), 0.1 mM luciferin (Promega, Madison, WI), and antibiotics (25 U/ml penicillin, 25 ug/ml streptomycin) (Yamazaki,S. 2005). Peripheral tissues were cut by hand to 1mm x 1 mm and placed on squares of polypropylene mesh (Spectramesh, Cole-Parmer, Chicago, IL) Dishes were sealed with circular glass coverslips and silicon grease. Cultures were maintained at 35.8° C and bioluminescence was measured using photomultiplier tubes (Hamamatsu, as supplied by Actimetrics Lumicycle System, Evanston, IL). Behavioral Analysis: Entrainment to the T cycle was assessed by stability of phase angle, and by a chi-square periodogram on the final 10 days of LD. Only animals with a stable phase angle and one dominant peak in the periodogram with a period of the LD cycle were used in subsequent studies. Free-running period in DD was assessed using a regression line fit to the three activity onsets in DD. Onsets were selected by the ClockLab software in most instances, but when the software did not appear to select an appropriate onset, two independent observers calculated the FRP, and in these cases the estimates were within 0.1h In vitro Analysis: The first day after dissection was not included due to transient activity. Once tissues were stabilized, up to a week of data was used, with a minimum of three days for any tissue. Baseline drift was subtracted from the data using a polynomial curve, of an order of one less than the number of days used, which was fit to the data. Using the baseline-subtracted data, we estimated circadian period by fitting a sine wave with an exponential decay (C=Asin(2π/P-t0)*e-t/τ, where A is the amplitude, P is the period, t0 is the phase, and tau is the damping constant). We only included data with a greater than 90% goodness of fit, as measured by the % variance accounted by the fit curve, with a median filter smoother of 6. Phase was determined from the peak measured on the second day in vitro, using a median smoother of 15 or 30. Data above from mouse 236 T21 T26 Constant Light (LL) • Conclusions • The SCN does express period aftereffects. Following a short T cycle, when FRP is shortened in vivo, SCN FRP is lengthened in vitro. This finding extends that of previous work with per1-luc mice [2]. This paradoxical result challenges our current understanding of the relationship of SCN circadian period in vitro and control of behavioral rhythms by the SCN in vivo. • The peripheral oscillators we investigated here (lung, thymus, spleen and esophagus) do not appear to express aftereffects in circadian period. • Phase relation of spleen and thymus to SCN is altered by T cycle pretreatment. This might be related to the well-established result that phase relation of the behavior to the light cycle is altered under different T cycles. • Our preliminary data suggest that LL lengthens behavioral FRP in vivo, but does not appreciably alter SCN FRP in vitro. Persistence of rhythmicity in vitro may be lessened by LL pretreatment. LL housed mice expressed lengthened behavioral FRP and less robust SCN FRP SCN SCN Raw Raw Baseline subtracted Baseline subtracted Raw SCN data from T21 mice for comparison Raw SCN data from LL mice This mouse showed a behavioral FRP of 22.95 h, and an SCN FRP of 24.35 h. This mouse showed a behavioral FRP of 24.46 h and an SCN FRP of 22.95 h. We would like to acknowledge financial support from NSF 0423200 and NSF RUI 0618129, Smith College Dean of the Faculty, Psychology Department funds. We thank Tanya Leise, Judy McKinley Brewer and Eric Bittman for helpful discussions, Qingsi Zhu, Natalie Strokes, and Caroline Walsh for technical assistance. Kate Dahlgren and Qingsi Zhu were supported in part by a summer stipend from HHMI.