Download

1 / 43
430 likes | 611 Views
Lehninger Ch. 19,23 BIO 322 Recitation 5 / Spring 2013. Oxidative Phosphorylation Lipid and Protein Biosynthesis Integration. Site of oxidative phosphorylation Mitochondria is like gram negative bacteria. Two membranes, Outer – permeable to small molecules via porin (transmembrane channels)

E N D
Lehninger Ch. 19,23 BIO 322 Recitation 5 / Spring 2013 Oxidative PhosphorylationLipid and Protein BiosynthesisIntegration
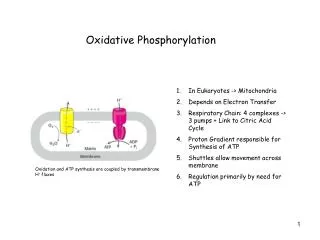
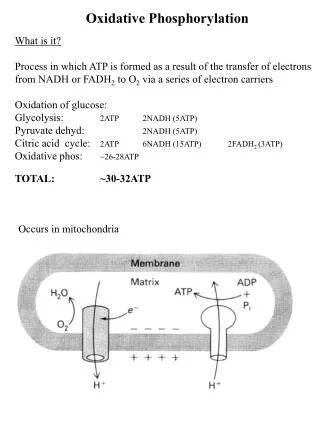
Site of oxidative phosphorylation • Mitochondria is like gram negative bacteria. Two membranes, • Outer – permeable to small molecules via porin (transmembrane channels) • Innermembrane is impermeable to most small molecules and ions, including protons (the only species that cross this membrane do so through specific transporters) – • Contains the components of respiratory chain and ATP synthase. • Matrix – enclosed by innermembrane, contains the PDH complex, TCA cycle enyzmes, beta oxidation enzymes and amino acid oxidation enzymes • Specific transporters carry pyruvate, fatty acids and their alpha keto derivatives into the matrix MITOCHONDRIA
Electron acceptors • Oxid. Phosphorylation begins with the entry of electrons into the respiratory chain. • Most of these electrons arise from action of dehyrogenases that collect electrons from catabolic pathways and funnel them to NAD, NADP and FAD • NAD-linked dehydrogenases remove two hydrogen atoms from their substrates. One to NAD as hydride ion (minus charged) and the other one to medium (plus charged) • Generally NADH carries electrons from catabolic rxns to respiratory chain • NADPH generally supplies electrons to anabolic rxns. • Reduced form/oxidized form relatively high for NADPH, low for NADH • NADH and NADPH cannot cross inner mitoch. membrane but electrons they carry can be shuttled across indirectly. • FAD can accept one (quinone to hydroquinone (semiquinone form)) or two electrons (dehydrogenations) – Can act as an intermediate. Unlike NAD and NADPH, the relevant standard reduction potential of FAD is dependent on the protein that it is associated with, not isolated FAD.
Membrane bound carriers • Three types of electron transfer occurs: 1) direct transfer of electrons Fe2+ to Fe3+ • 2) transfer as a hydrogen atom (H+) + electron • 3) transfer as a hydride ion (H-) • Reducing equivalent is used to designate a single electron equivalent transferred in an oxidation-reduction rxn. • In addition to NAD and flavoproteins, three other electron carrying molecules function in the respiratory chain. hydrophobic quinone (ubiquinone) and two different types of iron containing proteins – cytochromes and iron-sulfur proteins • Ubiquinone (also called coenzyme Q) • Can accept one or two electrons • Like flavoproteins can act at a junction between two • electron donor and a one-electron acceptor. • Lipid soluble, can freely diffuse within the lipid bilayer of • inner mitochondrial membrane. • Can carry both electrons and protons, central role in • Coupling electron flow to proton movement
Cytochromes, strong absorption of visible light due to their iron containing-heme prosthetic groups • Three classes – according to their light absorption spectra, a,b,c • Three absorption bands in reduced state. (longest wavelenth band - 600 nm for a, 560 nm for c, 550 nm for c) • Wavelent is used to distinguish between same class (absorption maximum). • Standard reduction potential of heme depends on its interaction with protein side chains. • Heme of cyt c – covalently bound, cyt a and b not covalent • Cyt a and b and some type of c are integral proteins of the inner mitochondrial memb. • Cyt c is soluble and associates through electrostatic interactions with outer surface of the inner membrane.
Iron sulfur proteins – iron is present not in heme but in association with inorganic sulfur atoms of Cys residue • One electron transfer, standard reduction potential dependent on microenvironment around iron. • Overall Rxn of mitoch. respiratory chain – electrons move from NADH, succinate through flavoproteins, ubiquinone, iron sulfur proteins, cytochromes and oxygen.
In the order of increasing reduction potential beucase electrons tend to flow from lower potential to higher potential. • NADH – Q- cyc b – cyc c1 – cyc c – cyc a – cyc a3 – oxygen • Not in this order in cellular conditions because actual reduction potential is dependent on concentration of reduced and oxidized forms.
Multienzyme complexes • 4 multi-enzyme complexes at mitoch. inner membrane. • Complex I and II transfer electron to ubiquinone from two different electron donors. NADH for complex I and succinate for complex II. • Complex III carries electrons from reduced ubiquinone to cyc c and complex IV completes the sequence by transferring electrons from cyc c to oxygen. • Complex I – NADH to Ubiquinone Oxidoreductase • or NADH deydrogenase • Catalyzes two simultaneous and coupled processes. • 1)Exergonic transfer to ubiquinone of a hydride ion from • NADH and a proton from the matrix • 2)Endergonic transfer of four protons from the matrix to • intermembrane
Complex 1 is a proton pump driven by the energy of electron transfer • The reaction it catalyzes is vectorial • It moves protons from matrix, which becomes negatively charged with the departure of protons, to the intermembrane space which becomes positively charged • Ubiquinol –QH2 diffuses in the inner mitochondrial membrane from complex I to complex III, where it is oxidized to Q in a process that also involves the outward movement of H+.
Complex II: Succinate to Ubiquinone • Succinate dehydrogenase, only membrane bound enzyme of the TCA cycle – CH 16. • Path of electron transfer: succinate binding site to FAD then through Fe-S center to the Q binding site. – 40 A long • But none of the electron transfer ditances exceeds 11 A – reasonable length for electron transfer. • Heme b of Complex II is apperantly not in the direct path of electron transfer – reduced frequency of electron leak, protects against ROS • Leak out of the system from succinate to oxygen – reason for ROS (hydrogen peroxide and superoxide radical) – SNP in this enyzme causes paraganglioma, benign tumors of head and neck commonly in carotid body, the organ that senses oxygen levels in blood.
Not through complex II • The first step of beta oxidation of fattyl acyl-CoA, catalyzed by flavoprotein acyl-CoA dehydrogenase. Transfer to FAD then to ETF (electron transfer flavorproteins), then to ETF:ubiquinone oxidoreductase – ubiquinone. • Glycerol-3 phosphate formed either from glycerol released by triacylglycerol breakdown or by reduction of (dihydroxyacetone P) DHAP from glycolysis is oxidized by glycerol 3-phosphate dehydrogenase. • This enzyme is a flavoprotein located on the outer face of the inner mitochondrial membrane, reduces ubiquinone.
Complex III:Ubiquinone to Cytochrome c • Cytochrome bc1 complex or ubiquinone:cytochrome c oxidoreductase • Couples from ubiquinol to cytochrome c with vectorial transport of protons from matrix to intermembrane space • Functional unit is a dimer with two monomeric units of cytochrome b surrounding a cavern in the middle of the membrane. • Ubiquinone is free to move from the matrix side of the membrane to the intermembrane space
Based on the structure of complex III – Q cycle • Q cycle accomodates the switch between the two electron carrier ubiquinone and one electron carriers – cytochrome b562,b566,c1 and c – 4 pair of electrons are translocated per pair of electrons through complex III to cytochrome c. • QH2 is oxidized to Q and two molecules of cyt c are reduced • Cyc c is a soluble protein of the intermembrane space to complex IV
Complex IV : cytochrome c oxidase • From cyc c to oxygen and reducing it to water • Electron transfer through complex IV is from cyc c to CuA center to heme a to the heme a3-cub center and finally to molecular oxygen • For every 4 electrons, consumes 4 substrate H from matrix • To avoid ROS formation, the intermediates remain tightly bound to the complex until completely converted to water.
Chemical potential energy Electrical potential energy
The passage of electrons from QH2 to cytochrome bl through complex III, and passage of electrons from complex I to QH2, involve the Q radical. Low probability pass an electron to O2 Superoxide free radical very reactive To prevent oxidative damage, cells have several forms of the enzyme superoxide dismutase, generates hydrogen peroxide. H2O2 to H2O by the action of glutathione peroxidase. Reduced GSH keeps protein sulfhydryl groups in their reduced state Recycled by PPP
CHEMIOSMOTIC MODEL The inner mitochondrial membrane is impermeable to protons. The proton motive force that drives the protons back into matrix, provides the energy for ATP synthesis
F1: a peripheral membrane protein • F0: integral to the membrane • ATP synthase catalyzes the formation of ATP from ADP and Pi, accompanied by the flow of protons. F0F1 binds ATP with very high affinity. Binding energy drives the eq toward formation.
Rotational catalysis 3 active sites of F1 take turns catalyzing ATP synthesis.
Beta subunit starts beta-ADP conformation to beta-ATP form, final beta-empty conformation
This antiporter moves four negative charges out for every three moved in. Its activity is favored by the transmembrane electrochemical gradient gives the matrix net negative charge
Inner membrane is not permeable to NADH. How can NADH generated in the cytosol be reoxidized to NAD+ by oxygen via respiratory chain? Liver, kidney, heart NADH to respiratory chain 2.5 ATP per pair of electron.
Skeletal muscle and brain use a different NADH shuttle. • It delivers the reducing equivalents from NADH to Q Complex III – not complex I • Providing enough energy to synthesize 1.5 ATP per pair of electrons.
ATP PRODUCING PATHWAYS ARE COORDINATELY REGULATED ATP consumption increases rate of ETC increases When conversion of ADP to ATP lowers the ADP concentration, electron transfer slows, glycolysis and TCA slows ATP is an allosteric inhibitor of PFK-1 and pyruvate dehydrogenase