Download
1 / 28
280 likes | 492 Views
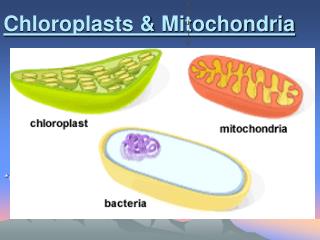
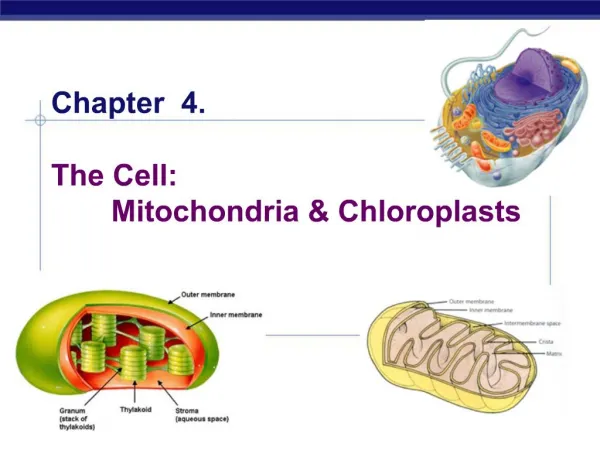
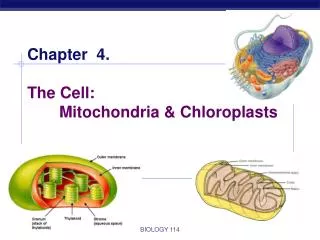
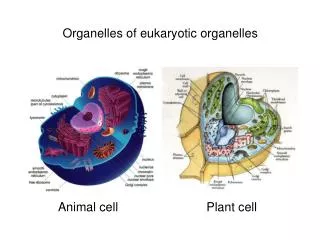
Energy production in eukaryotic organelles Mitochondria and chloroplasts. Energy production in eukaryotic organelles Mitochondria. Source of energy Glycolysis and citric acid cycle. Three stages:

E N D
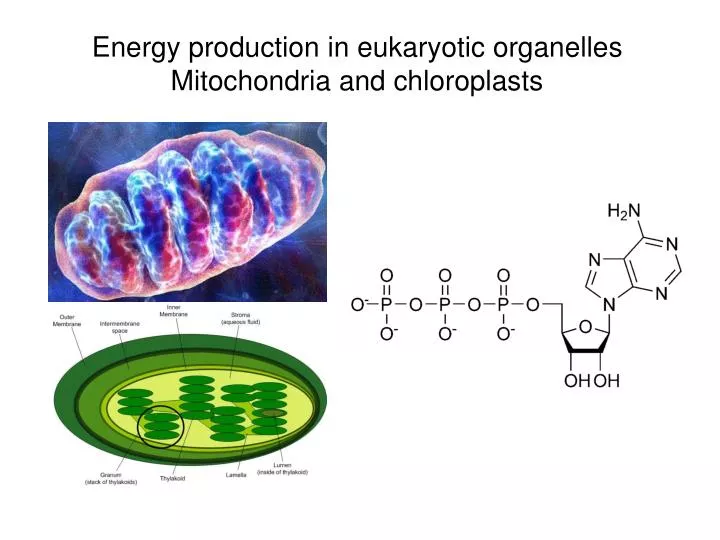
Energy production in eukaryotic organellesMitochondria and chloroplasts
Source of energyGlycolysis and citric acid cycle Three stages: • glycolysis converts glucose into 2 three carbon molecules (pyruvate) PLUS (substrate level phosphorylation of ATP) • citric acid cycle breaks down glucose into CO2 • electron transport chain passes electrons down a chain to eventually bond with hydrogen molecules to form water (part of chemiosmosis)
NAD+, NADH; FAD, FADH2 NAD+ FAD FAD reduced to FADH2,
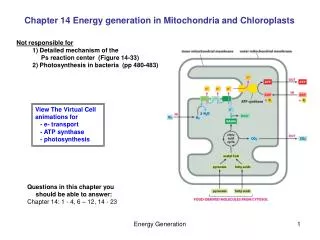
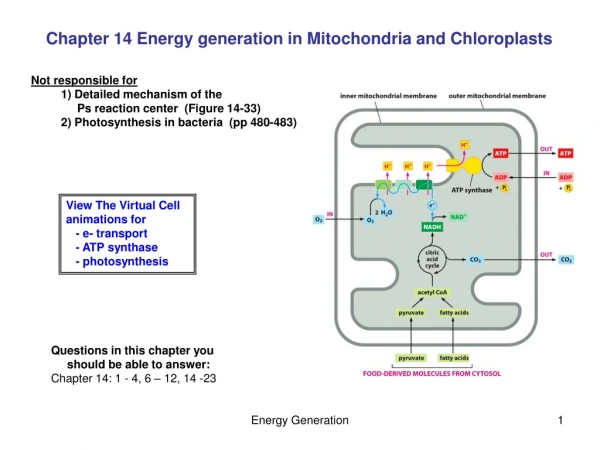
Respiratory chain in mitochondria • The cells of all eukaryotes (all animals, plants, fungi, algae, protozoa – in other words, all living things except bacteria and archaea) contain intracellular organelles called mitochondria, which produce ATP. Energy sources such as glucose are initially metabolized in the cytoplasm. The products are imported into mitochondria. Mitochondria continue the process of catabolism using metabolic pathways including the Krebs cycle, fatty acid oxidation, and amino acid oxidation. • The end result of these pathways is the production of two kinds of energy-rich electron donors, NADH and FADH2. Electrons from these donors are passed through an electron transport chain to oxygen, which is reduced to water. This is a multi-step redox process that occurs on the mitochondrial inner membrane. The enzymes that catalyze these reactions have the remarkable ability to simultaneously create a proton gradient across the membrane, producing a thermodynamically unlikely high-energy state with the potential to do work.
Components of the mitochondrial electron transport chain • Complex IComplex I (NADH dehydrogenase, also called NADH:ubiquinone oxidoreductase) removes two electrons from NADH and transfers them to a lipid-soluble carrier, ubiquinone (Q). • Complex IIComplex II (succinate dehydrogenase) serves to funnel additional electrons into the quinone pool (Q) by removing electrons from succinate and transferring them (via FAD) to Q. • Complex IIIComplex III (cytochrome bc1 complex) removes in a stepwise fashion two electrons from QH2 and transfers them to two molecules of cytochrome c • Complex IVComplex IV (cytochrome c oxidase) removes four electrons from four molecules of cytochrome c and transfers them to molecular oxygen (O2) http://www.elmhurst.edu/~chm/vchembook/596electransport.html
Respiratory chain • Complex I • Complex I (NADH dehydrogenase, also called NADH:ubiquinone oxidoreductase; EC 1.6.5.3) removes two electrons from NADH and transfers them to a lipid-soluble carrier, ubiquinone (Q). The reduced product, ubiquinol (QH2) is free to diffuse within the membrane. At the same time, Complex I moves four protons (H+) across the membrane, producing a proton gradient. Complex I is one of the main sites at which premature electron leakage to oxygen occurs, thus being one of main sites of production of a harmful free radical called superoxide. • Complex II • Complex II (succinate dehydrogenase; EC 1.3.5.1) is not a proton pump. It serves to funnel additional electrons into the quinone pool (Q) by removing electrons from succinate and transferring them (via FAD) to Q. Complex II consists of four protein subunits: SDHA,SDHB,SDHC, and SDHD. Other electron donors (e.g., fatty acids and glycerol 3-phosphate) also funnel electrons into Q (via FAD), again without producing a proton gradient. • Complex III • Complex III (cytochrome bc1 complex; EC 1.10.2.2) removes in a stepwise fashion two electrons from QH2 and transfers them to two molecules of cytochrome c, a water-soluble electron carrier located within the intermembrane space. At the same time, it moves two protons across the membrane, producing a proton gradient (in total 4 protons: 2 protons are translocated and 2 protons are released from ubiquinol). When electron transfer is hindered (by a high membrane potential, point mutations or respiratory inhibitors such as antimycin A), Complex III may leak electrons to oxygen resulting in the formation of superoxide, a highly-toxic species, which is thought to contribute to the pathology of a number of diseases, including aging. • Complex IV • Complex IV (cytochrome c oxidase; EC 1.9.3.1) removes four electrons from four molecules of cytochrome c and transfers them to molecular oxygen (O2), producing two molecules of water (H2O). At the same time, it moves four protons across the membrane, producing a proton gradient. • The pathway of electrons occurs as follows: NADH is oxidized to NAD+, reducing Flavin mononucleotide to FMNH2 in one two-electron step. The next electron carrier is a Fe-S cluster, which can only accept one electron at a time to reduce the ferric ion into a ferrous ion. In a convenient manner, FMNH2 can be oxidized in only two one-electron steps, through a semiquinone intermediate. The electron thus travels from the FMNH2 to the Fe-S cluster, then from the Fe-S cluster to the oxidized Q to give the free-radical (semiquinone) form of Q. This happens again to reduce the semiquinone form to the ubiquinol form, QH2. During this process, four protons are translocated across the inner mitochondrial membrane, from the matrix to the intermembrane space. This creates a proton gradient that will be later used to generate ATP through oxidative phosphorylation.
Cytochromes • Cytochromes are, in general, membrane-bound hemoproteins that contain heme groups and carry out electron transport. • They are found either as monomeric proteins (i.e., cytochrome c) or as subunits of bigger enzymatic complexes that catalyze redox reactions. • The heme group is a highly-conjugated ring system (which means its electrons are very mobile) surrounding a metal ion, which readily interconverts between the oxidation states. For many cytochromes, the metal ion present is that of iron, which interconverts between Fe2+ (reduced) and Fe3+ (oxidized) states (electron-transfer processes) or between Fe2+ (reduced) and Fe3+ (formal, oxidized) states (oxidative processes). Cytochromes are, thus, capable of performing oxidation and reduction. http://www.rpi.edu/dept/bcbp/molbiochem/MBWeb/mb1/part2/redox.htm
Electron flow in mitochondria Stepwise flow of electrons through the electron transport chain from NADH, succinate, and FADH2 to O2 (blue arrows). Each of the four large multi-protein complexes in the chain is located in the inner mitochondrial membrane and contains several specific electron carriers. Coenzyme Q (CoQ) and cytochrome c transport electrons between the complexes. As shown by the redox scale, electrons pass in sequence from carriers with a lower reduction potential to those with a higher (more positive) potential. The free-energy scale shows the corres-ponding reduction in free energy as a pair of electrons moves through the chain. The energy released as electrons flow through three of the complexes is sufficient to power the pumping of H+ ions across the membrane, establishing a proton-motive force.
ATP production by oxidative phosphorylation • The chemiosmotic coupling hypothesis explains that the electron transport chain and oxidative phosphorylation are coupled by a proton gradient across the inner mitochondrial membrane. The efflux of protons creates both a pH gradient and an electrochemical gradient. This proton gradient is used by the FOF1 ATP synthase complex to make ATP via oxidative phosphorylation. ATP synthase is sometimes regarded as complex V of the electron transport chain. The FO component of ATP synthase acts as an ion channel for return of protons back to mitochondrial matrix. During their return, the free energy produced during the generation of the oxidized forms of the electron carriers (NAD+ and FAD) is released. This energy is used to drive ATP synthesis, catalyzed by the F1 component of the complex. • Coupling with oxidative phosphorylation is a key step for ATP production. However, in certain cases, uncoupling may be biologically useful. The inner mitochondrial membrane of brown adipose tissue contains a large amount of thermogenin (an uncoupling protein), which acts as uncoupler by forming an alternative pathway for the flow of protons back to matrix. This results in consumption of energy in thermogenesis rather than ATP production. This may be useful in cases when heat production is required, for example in colds or during arise of hibernating animals. Synthetic uncouplers (e.g., 2,4-dinitrophenol) also exist, and, at high doses, are lethal.
ATPase • An ATP synthase is a general term for an enzyme that can synthesize adenosine triphosphate (ATP) from adenosine diphosphate (ADP) and inorganic phosphate by using some form of energy. This energy is often in the form of protons moving down a electrochemical gradient, such as from the lumen into the stroma of chloroplasts or from the inter-membrane space into the matrix in mitochondria. The overall reaction sequence is: • ADP + Pi → ATP • These enzymes are of crucial importance in almost all organisms, because ATP is the common "energy currency" of cells. • The antibiotic oligomycin inhibits the FO unit of ATP synthase.
Mechanism of ATP production The binding change mechanism involves the active site of a β subunit cycling between three states. In the "open" state, ADP and phosphate enter the active site, in the diagram to the right this is shown in brown. The protein then closes up around the molecules and binds them loosely - the "loose" state (shown in red). The enzyme then undergoes another change in shape and forces these molecules together, with the active site in the resulting "tight" state (shown in pink) binding the newly-produced ATP molecule with very high affinity. Finally, the active site cycles back to the open state, releasing ATP and binding more ADP and phosphate, ready for the next cycle of ATP production.
A proton gradient is the driving force for ATP production Key points: 1. Protons are translocated across the membrane, from the matrix to the intermembrane space, as a result of electron transport resulting from the formation of NADH by oxidation reactions. (See the animation of electron transport.) The continued buildup of these protons creates a proton gradient. 2. ATP synthase is a large protein complex with a proton channel that allows re-entry of protons. 3. ATP synthesis is driven by the resulting current of protons flowing through the membrane http://www.sp.uconn.edu/~terry/images/anim/ATPmito.html
Energy production in eukaryotic organellesChloroplasts http://www.helpsavetheclimate.com/photosynthesis.html
Light for photosynthesis • What we call "light" is actually part of the spectrum of electromagnetic radiation. This covers a range of rays or waves from cosmic rays, with very short wavelengths all the way to radio waves
Light harvesting • The absorption of light in chloroplasts is by a range of pigments, mainly chlorophylls (which provide the green colour) and carotenoids. These different types of pigment absorb light of different wavelengths. A large number of these pigment molecules are arranged into an antenna to capture most of the photons of the correct energy. This energy is then passed through a chain of pigment molecules until it reaches a specialised molecular complex called the reaction centre. The pigment antenna complex/ reaction centre together are called a photosystem
Chemical structure of chlorophyll • Chlorophyll is a chlorin pigment, which is structurally similar to and produced through the same metabolic pathway as other porphyrin pigments such as heme. At the center of the chlorin ring is a magnesium ion. The chlorin ring can have several different side chains, usually including a long phytol chain. There are a few different forms that occur naturally, but the most widely distributed form in terrestrial plants is chlorophyll a. The general structure of chlorophyll a was elucidated by Hans Fischer in 1940, and by 1960, when most of the stereochemistry of chlorophyll a was known • The different types of chlorophyll are Chlorophyll a, Chlorophyll b, Chlorophyll c1, Chlorophyll c2 and Chlorophyll d. These types vary in side chains.
Electron Transport Pathway from Water (to NADP+ (Nicotinamide Adenine Dinucleotide Phosphate, oxidized form) Abbreviations used are (from left to the right of the diagram): Mn for a manganese complex containing 4 Mn atoms, bound to Photosystem II (PSII) reaction center; O2 for oxygen; H+ for protons; P680 for the reaction center chlorophyll (Chl) in PSII: it is the primary electron donor of PSII; Excited (Chl) P680 for P680* that has the energy of the photon of light; Pheo for pheophytin molecule (the primary electron acceptor of PSII; it is like a chlorophyll a molecule where magnesium (in its center) has been replaced by two "H"s); QA for a plastoquinone molecule tightly bound to PSII; QB for another plastoquinone molecule that is loosely bound to PSII; FeS for Rieske Iron Sulfur protein; Cyt. f for Cytochrome f; Cytb6 (L and H) for Cytochrome b6 (of Low and High Energy); PC for copper protein plastocyanin; P700 for the reaction center chlorophyll (Chl;actually a dimer, i.e., two molecules together) of PSI; it is the primary electron donor of PSI; Excited (Chl) P700 for P700* that has the energy of the photon of light; Ao for a special chlorophyll a molecule (primary electron acceptor of PSI); A1 for a phylloquinone (Vitamin K) molecule; FX, FA, and FB are three separate Iron Sulfur Centers; FD for ferredoxin; and FNR for Ferredoxin NADP oxido Reductase (FNR). Three major protein complexes are involved in running the "Z" scheme: (1) Photosystem II; (2) Cytochrome bf complex (containing Cytb6; FeS; and Cytf ) and (3) Photosystem I. Z-scheme http://web.virginia.edu/Heidi/chapter22/chp22.htm
Cyclic and non cyclic photophosphorylation • Cyclic photophosphorylation • In cyclic electron flow, the electron begins in a pigment complex called photosystem I, passes from the primary acceptor to ferredoxin, then to a complex of two cytochromes (similar to those found in mitochondria), and then to plastoquinone before returning to chlorophyll. This transport chain produces a proton-motive force, pumping H+ ions across the membrane; this produces a concentration gradient which can be used to power ATP synthase during chemiosmosis. This pathway is known as cyclic photophosphorylation, and it produces neither O2 nor NADPH. In bacterial photosynthesis, a single photosystem is used, and therefore is involved in cyclic photophosphorylation. Noncyclic photophosphorylation The other pathway, noncyclic photophosphorylation, is a two-stage process involving two different chlorophyll photosystems. First, a water molecule is broken down into 2H+ + 1/2O2 + 2e- by a process called photolysis (or light-splitting). The two electrons from the water molecule are kept in photosystem II, while the 2H+ and 1/2O2 are left out for further use. Then a photon is absorbed by the chlorophyll core of photosystem II, exciting the two electrons which are transferred to the acceptor molecule. The deficit of electrons is replenished by taking electrons from another molecule of water. The electrons transfer from the primary acceptor to plastoquinone, then to plastocyanin, producing proton-motive force as with cyclic electron flow and driving ATP synthesis. The photosystem II complex replaced its lost electrons from an external source, however, the two other electrons are not returned to photosystem II as they would in the analogous cyclic pathway. Instead, the still-excited electrons are transferred to a photosystem I complex, which boosts their energy level to a higher level using a second solar photon. The highly excited electrons are transferred to the acceptor molecule, but this time are passed on to an enzyme called Ferredoxin- NADP reductase|NADP+ reductase, for short FNR, which uses them to catalyst the reaction (as shown): NADP+ + 2H+ + 2e- → NADPH + H+ This consumes the H+ ions produced by the splitting of water, leading to a net production of 1/2O2, ATP, and NADPH+H+ with the consumption of solar photons and water. The concentration of NADPH in the chloroplast may help regulate which pathway electrons take through the light reactions. When the chloroplast runs low on ATP for the Calvin cycle, NADPH will accumulate and the plant may shift from noncyclic to cyclic electron flow.
Electron transport in the chloroplast membraneATP production by H+ gradient
Chloroplast ATP synthase (ATPase) • In plants ATP synthase is present in chloroplasts (CF1FO-ATP synthase). The enzyme is integrated into thylakoid membrane; the CF1-part sticks into stroma, where dark reactions of photosynthesis (Also called the light-independent reactions or the Calvin cycle) and ATP synthesis take place. The overall structure and the catalytic mechanism of the chloroplast ATP synthase are almost the same as those of the mitochondrial enzyme. However, in chloroplasts the proton motive force is generated not by respiratory electron transport chain, but by primary photosynthetic proteins.
Light-independent reaction In photosynthesis, the light-independent reactions, also somewhat misleadingly called the dark reactions (they don't require darkness, but they require the products of the light reactions), are chemical reactions that convert carbon dioxide and other compounds into glucose. It occurs in the stroma, the fluid filled area of a chloroplast outside of the thylakoid membranes. These reactions, unlike the light-dependent reactions, do not need light to occur; hence the term dark reactions. These reactions take the products of the light-dependent reactions and perform further chemical processes on them. There are three phases to the light-independent reactions, collectively called the Calvin Cycle: Carbon Fixation, Reduction reactions, and ribulose 1,5-biphosphate (RuBP) regeneration.
The Calvin cycle • The enzymes in the Calvin cycle are functionally equivalent to many enzymes used in other metabolic pathways such as gluconeogenesis and the pentose phosphate pathway, but they are to be found in the chloroplast stroma instead of the cell cytoplasm, separating the reactions. They are activated in the light (which is why the name "dark reaction" is misleading), and also by products of the light-dependent reaction. These regulatory functions prevent the Calvin cycle from operating in reverse to respiration, which would create a continuous cycle of carbon dioxide being reduced to carbohydrates, and carbohydrates being respired to carbon dioxide. Energy (in the form of ATP) would be wasted in carrying out these reactions that have no net productivity. • The sum of reactions in the Calvin cycle is the following: • 3 CO2 + 6 NADPH + 5 H2O + 9 ATP → C3H5O3-PO32- + 2 H+ + 6 NADP+ + 9 ADP + 8 PiOR3 CO2 + 6 C21H29N7O17P3 + 5 H2O + 9 C10H16N5O13P3 → C3H5O3-PO32- + 2 H+ + 6 NADP+ + 9 C10H15N5O10P2 + 8 Pi • It should be noted that hexose (six carbon) sugars are not a product of the Calvin cycle. Although many texts list a product of photosynthesis as C6H12O6, this is mainly a convenience to counter the equation of respiration, where six-carbon sugars are oxidized in mitochondria. The carbohydrate products of the Calvin Cycle are three-carbon sugar phosphate molecules, or "triose phosphates," specifically, glyceraldehyde-3-phosphate.
Steps of the Calvin cycle • The enzyme RuBisCO catalyses the carboxylation of Ribulose-1,5-bisphosphate, a 5 carbon compound, by carbon dioxide (a total of 6 carbons) in a two-step reaction [1]. Rubisco is a large, slow enzyme averaging 3 substrate per second compared to 1000/s for most other enzymes in the Calvin cycle. Two molecules of glycerate 3-phosphate, a 3-carbon compound, are created. (also: 3-phosphoglycerate, 3-phosphoglyceric acid, 3PGA) • The enzyme phosphoglycerate kinase catalyses the phosphorylation of 3PGA by ATP (which was produced in the light-dependent stage). 1,3-bisphosphoglycerate (glycerate-1,3-bisphosphate) and ADP are the products. (However, note that two PGAs are produced for every CO2 that enters the cycle, so this step utilizes 2ATP per CO2 fixed. • The enzyme G3P dehydrogenase catalyses the reduction of 1,3BPGA by NADPH (which is another product of the light-dependent stage). Glyceraldehyde 3-phosphate (also G3P, GP) is produced, and the NADPH itself was oxidized and becomes NADP+. Again, two NADPH are utilized per CO2 fixed. • (Simplified versions of the Calvin cycle integrate the remaining steps, except for the last one, into one general step - the regeneration of RuBP - also, one G3P would exit here.) • Triose phosphate isomerase converts some G3P reversibly into dihydroxyacetone phosphate (DHAP), also a 3-carbon molecule. • Aldolase and fructose-1,6-bisphosphatase convert a G3P and a DHAP into fructose-6-phosphate (6C). A phosphate ion is lost into solution. • Then fixation of another CO2 generates two more G3P. • F6P has two carbons removed by transketolase, giving erythrose-4-phosphate. The two carbons on transketolase are added to a G3P, giving the ketose xylulose-5-phosphate (Xu5P). • E4P and a DHAP (formed from one of the G3P from the second CO2 fixation) are converted into sedoheptulose-1,7-bisphosphate (7C) by aldolase enzyme. • Sedoheptulose-1,7-bisphosphatase (one of only three enzymes of the Calvin cycle which are unique to plants) cleaves sedoheptulose-1,7-bisphosphate into sedoheptulose-7-phosphate, releasing an inorganic phosphate ion into solution. • Fixation of a third CO2 generates two more G3P. The ketose S7P has two carbons removed by transketolase, giving ribose-5-phosphate (R5P), and the two carbons remaining on transketolase are transferred to one of the G3P, giving another Xu5P. This leaves one G3P as the product of fixation of 3 CO2, with generation of three pentoses which can be converted to Ru5P. • R5P is converted into ribulose-5-phosphate (Ru5P, RuP) by phosphopentose isomerase. Xu5P is converted into RuP by phosphopentose epimerase. • Finally, phosphoribulokinase (another plant unique enzyme of the pathway) phosphorylates RuP into RuBP, ribulose-1,5-bisphosphate, completing the Calvin cycle. This requires the input of one ATP. • Thus, of 6 G3P produced, three RuBP (5C) are made totalling 15 carbons, with only one available for subsequent conversion to hexose. This required 9 ATPs and 6 NADPH per 3 CO2. • RuBisCO also reacts competitively with O2 instead of CO2 in photorespiration. The rate of photorespiration is higher at high temperatures. "photorespiration" turns RuBP into 3PGA and 2-phosphoglycolate, a 2-carbon molecule which can be converted via glycolate and glyoxalate to glycine. Via the glycine cleavage system and tetrahydrofolate, two glycines are converted into serine +CO2. Serine can be converted back to 3-phosphoglycerate. Thus, only 3 of 4 carbons from two phosphoglycolates can be converted back to 3PGA. Obviously photorespiration has very negative consequences for the plant, because rather than fixing CO2), this process leads to loss of CO2). C4 carbon fixation evolved to circumvent photorespiration, but can only occur in certain plants living in very warm or tropical climates.
Substrate level phosphorylation • Glycolysis is the sequence of reactions that converts glucose into pyruvate with the concomitant production of a relatively small amount of adenosine triphosphate (ATP). The word is derived from Greek γλυκύς (sweet) and λύσις (letting loose). • It is the initial process of most carbohydrate catabolism, and it serves three principal functions: • 1. Generation of high-energy molecules (ATP and NADH) as cellular energy sources as part of aerobic respiration and anaerobic respiration; that is, in the former process, oxygen is present, and, in the latter, oxygen is not present • 2. Production of pyruvate for the citric acid cycle as part of aerobic respiration • 3. Production of a variety of six- and three-carbon intermediate compounds, which may be removed at various steps in the process for other cellular purposes.