Download

1 / 122
1.23k likes | 1.38k Views
第六章 植物生长调节物质 植物体内合成的,以痕量水平存在,能对植物生理过程产生影响和调节作用的天然有机化合物称 植物激素 ( plant hormones )。 有 3 个特点 : 1 )内源的; 2 )能移动的; 3 )有调节功能 ; 是一种信号分子。我们把一些人工合成的具有天然植物激素相似生理功能的物质称植物生长调节剂,二者合称植物生长调节物质。

E N D
第六章 植物生长调节物质 植物体内合成的,以痕量水平存在,能对植物生理过程产生影响和调节作用的天然有机化合物称植物激素(plant hormones)。 有3个特点:1)内源的;2)能移动的;3)有调节功能;是一种信号分子。我们把一些人工合成的具有天然植物激素相似生理功能的物质称植物生长调节剂,二者合称植物生长调节物质。 目前公认的植物激素有5大类:IAA、GA、CTK、ABA、ETH。30年代开始研究IAA,50年代确定了GA和CTK,60年代又确定了ABA和ETH。正是这5大类植物激素的相互联系、相互作用调节着植物的细胞分裂、伸长,调节着植物的生长发育。
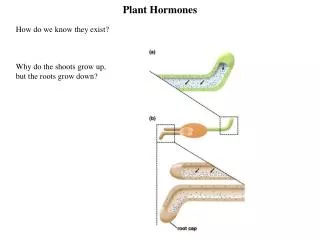
研究植物激素的基本方法为:1)外源施加法;2)内源分析法;3)使用抑制剂,包括合成、作用、运输抑制剂等;4)突变体研究,有缺失突变体如矮杆玉米缺GA和不敏感突变体,其对某种激素不敏感。5)转达基因植物研究。研究植物激素的基本方法为:1)外源施加法;2)内源分析法;3)使用抑制剂,包括合成、作用、运输抑制剂等;4)突变体研究,有缺失突变体如矮杆玉米缺GA和不敏感突变体,其对某种激素不敏感。5)转达基因植物研究。 第一节 生长素(Auxin) 一、生长素的发现 1、1880年英国Charles Darwin 和Francis Darwin,用单侧光照射金丝雀Yi草的胚芽鞘会发生向光性弯曲。指出在单侧光照射下,幼苗会产生某种影响从上部传到下部,造成向光面和背光面不等生长。 2、1913年丹麦 Boysen-Jensen,用燕麦胚芽鞘试验,证明Darwin所谓的影响可以透过凝胶块。
3、1914年和1919年,匈牙利人A Paal 证实,胚芽鞘的顶端扩散性化合物具有促进生长的效应。 4、1928年荷兰人F.W.Went将燕麦胚芽鞘尖端切下,放在琼胶薄片上,约1小时后,移去芽鞘尖端,再将琼胶切成小块后,放在去顶的胚芽鞘一侧,置于暗中,胚芽鞘会向放琼胶小块的对侧弯曲。证明,芽鞘尖端的影响可传到琼胶。F.W. Went将这种物质称为生长素。 5、1934年荷兰人F.Kogl 从人尿中分离并提纯到了能刺激植物生长的物质,经鉴定为吲哚乙酸。 6、直到1942年Haagen Smit等才从碱性水解的玉米粉和未成熟的玉米种子胚乳中分离得到了吲哚乙酸。
二、生长素分子结构、体内分布和传导 (一)生长素分子结构 1、内源生长素结构 1)吲哚乙酸:广泛存在于植物体中。 2)吲哚乙腈:1952年由Jone在十字花科植物中发现。 3)吲哚丁酸:普遍存在于植物中。 4)4-氯–3-吲哚乙酸:1963年由日本人Marumo在未成熟的 豌豆种子中提取到。 5)苯乙酸:存在玉米、大麦、向日葵、烟草、番茄等植物中 。
2、生长素类生长调节剂结构 1)吲哚类:吲哚丙酸(IPA) 2)萘环类:萘乙酸(NAA)和萘氧乙酸 3)苯环类: 2,4-二氯苯氧乙酸 (2,4-D) 2,4,6-三氯苯氧乙酸 (2,4,6-T) 4-碘苯氧乙酸 (增产灵)。
(二)植物体内分布 广泛分布于植物的根、茎、叶、花、果实、种子、胚芽鞘中。一般生长旺盛的组织如形成层、幼果、幼种子、芽含量高。量值:10-100 ng/gFW。 (三)植物体内的运输 发生在幼茎、幼根和胚芽等薄壁细胞之间,具极性运输性质。生长素只能从植物形态学的上端向植物形态学的下端运输称IAA的极性运输。 IAA极性运输的特点为: 1)只发生在薄壁细胞之间。 2)为主动运输,IAA可逆浓度梯度运输,且需要O2。 3)IAA极性运输速度比物理扩散大10倍。
IAA极性运输的机理: (化学渗透极性扩散学说)认为: 1)质膜上有H+-ATPase,水解ATP,不断将H+泵到细胞壁。胞间质pH较低,IAA常以-COOH形式存在(IAAH);胞内pH较高,IAA以-COO-形式存在(IAA-)。 2)IAAH易透过膜进入细胞内,形成IAA-,IAA-为亲水性的,不易透过膜。 3)仅细胞基部存在IAA-运输载体,造成IAA的极性运输。
IAA-运输载体已在植物中找到,发现该载体只存在于细胞有基部。IAA-运输载体已在植物中找到,发现该载体只存在于细胞有基部。 许多物质如TIBA(三碘苯甲酸)、NPA(萘基邻氨甲酰苯甲酸)可与IAA-运输载体结合,而使其构象发生变化,而抑制IAA的极性运输。这些物质称整形素或形态素。
茎尖和叶片合成的IAA可经维管束薄壁细胞, 沿维管束长距离运输,而不经韧皮部或木质部, 也有极性。速度约1~2.4 cm/h。 三、生长素的生物合成: (一)生物合成部位 根尖、茎尖、幼叶、正在发育的种子和果实,包括成熟叶片都能合成IAA。
(二)生长素合成途径: 1、依赖色氨酸的合成途径: 依据: 1)C14-Try饲喂给小麦、甜瓜叶片,分离得到了C14-IAA。 2)植物缺锌时,体内IAA含量会减少,得小叶病。因为Zn是色氨酸合成酶的组分,缺锌阻碍了吲哚和丝氨酸合成Try,加入锌,随着Try的合成,合成IAA的能力逐渐恢复。 1)吲哚丙酮酸途径:是高等植物合成IAA的主要途径。 Try转氨酶吲哚丙酮酸脱羧酶 3-吲哚乙醛氧化酶 Try 吲哚丙酮酸3-吲哚乙醛 IAA NH3 CO22H+ 3-吲哚乙醛氧化酶是一个关键酶。拟南芥有一种生长素超量积累突变体,称超级根(superroot),该突变体的3-吲哚乙醛氧化酶活性特别高。
2)吲哚乙醛肟途径: 3-吲哚乙腈水解酶 Try 吲哚3-乙醛肟3-吲哚乙腈 IAA 3-吲哚乙腈水解酶(nitrilase)是一个关键酶。十字花科、芭蕉等植物行此途径。最近,拟南芥得到了4种腈水解酶突变体:NIT1、NIT2、NIT3、NIT4。 3)色胺途径: Try 脱羧酶 胺氧化酶 3-吲哚乙醛氧化酶 Try 色胺3-吲哚乙醛 IAA CO2 NH3 2H+ 本途径在植物中占少数。大麦、燕麦、烟草和番茄在进行吲哚丙酮酸途时,还进行色胺途径合成IAA。
4)3-吲哚乙酰胺途径:仅发生在细菌中 在细菌中有一个Ti质粒,Ti质粒上有一个T-DNA,可随机插入植物体基因中进行表达。T-DNA上有2个基因控制IAA的合成,使感染组织的IAA增高。其中iaaM基因编码 Try单加氧酶,iaaH基因编码3-吲哚乙酰胺水解酶。 Try单加氧酶(IaaM) 3-吲哚乙酰胺水解酶(iaaH) Try 3-吲哚乙酰胺 IAA T-DNA还有一个基因,其控制IAA与赖氨酸结合, IAA与赖氨酸结合后形成吲哚3-乙酰赖氨酸,使IAA转化为结合态,从而降低IAA的含量。将iaaM基因转移到烟草中后,烟草体内的IAA含量是正常株的10倍,植物非常矮。
2、非色氨酸途径 Rapparini (1999)用15N-邻氨基苯甲酸和2H-Trp标记浮萍的复叶, 发现在起初的1h, IAA从15N-邻氨基苯甲酸结合的15N快于从2H-Trp 结合的2H,表明最初合成的IAA是Trp非依赖性的。在2h后, 2H-IAA才高于15N-IAA。 Normanly J, Bartel B. Redundancy as a way of life-IAA metabolism . Current Opinion in Plant Biology. 1999, 2: 207-213
四、生长素在植物体内的存在状态与降解 (一)束缚态和游离态生长素 1、束缚态生长素 植物体内的生长素可与细胞内的糖、氨基酸等物质结合而形成束缚态生长素。 如和葡萄糖形成:吲哚乙酰葡萄糖基酯,与ASP结合形成:吲哚乙酰天门冬氨酸。束缚态生长素需经水解、酶解或细胞自溶作用才能释放出,没有活性。其作用: 1)作为IAA的贮藏形式;2)作为IAA的运输形式; 3)解毒作用;4)防止IAA的氧化作用;5)调节自由IAA的含量。 2、游离型生长素:不与其它物质以共价键形式结合,易用甲醇、乙醚等有机溶剂从植物中提取出的生长素。其有活性。
(二)生长素在植物体内的降解:两条途径 1、 经IAA氧化酶降解:有脱羧和非脱羧降解两种(见下图): AA氧化酶早在1947年就被汤玉玮和Bonner 在豌豆幼茎的提取液中发现。其实是一个能引起IAA氧化分解的过氧化物酶。 IAA氧化酶分子中含铁血红蛋白,需Mn2+、和带一个羟基的一元酚如P-香豆酸、阿魏酸、2,4-二氯酚等作为辅助因子。单元酚抑制生长就这个原因。IAA氧化酶在体内的分布与植物的生长速率有关,一般生长旺盛的部位IAA氧化酶的含量比老组织中少,茎又比根中少。 2、光氧化: IAA极易发生光氧化,在有核黄素、紫黄素或合成色素存在下,IAA的光氧化大大加速。产物是吲哚醛和3-亚甲基羟吲哚。
五、生长素的生理作用 1、促进细胞伸长生长 IAA能促进植物茎切段和胚芽鞘切段的伸长生长。此外IAA还可促进马铃署和菊芋的块茎、组织培养中愈伤组织的生长。IAA对生长的作用有三个特点: 1)低浓度IAA促进生长,高浓度抑制生长。 2)不同器官对IAA的敏感性不同,对IAA的敏感性,根>芽>茎;不同年龄的细胞对IAA的敏感性也不同,幼龄>老龄;分化程度低的组织>分化程度高的组织;黄化组织>绿组织。 3)离体器官和整株植物对IAA反应不同:IAA对离体器官有明显的促进作用,而对整株植物效果不明显。
2、抑制侧芽生长,促进顶端优势 摘去顶芽的枝条,侧芽就萌发。除去顶芽后,以IAA的羊毛脂处理,侧芽就不萌发。推测顶芽合成的IAA抑制了侧芽的萌发。 机理: 认为顶芽合成的IAA经极性运输运到侧芽,由于侧芽对IAA较茎敏感,较高浓度的IAA就抑制了侧芽的生长;距顶芽越远,IAA氧化酶活性愈强,IAA氧化分解加强,IAA对侧芽的抑制作用也就减弱。
3、促进侧根和不定根的生长。 NAA+吲哚丁酸各50%构成生根粉。组织培养中控制培养基中IAA/CTK比例以调节根芽分化,比值大促进生根,反之利于长茎; IAA/GA比例调节维管束的分化方向,比值大利于木质部的分化,比值小利于韧皮部的分化。 4、向性生长 1)向光性:多数植物在单侧光照射下,背光面含较多IAA,发生 向光性弯曲。少数植物向光性还与生长抑制剂有关。 2)向重性:在重力的作用下,近地处IAA较高,由于根的敏感性较高,抑制近地面的生长。
5、促进瓜类雌花的分化和花粉的育性有关 用5-10ng/L的NAA或2,4-D处理营养生长达14个月的菠萝,2个月后就能开花。IAA处理可使黄瓜多开雌花。此外。花粉育性与IAA不足有关。 光敏不育水稻58S, 光照大于14小时/天:花粉败育。光照小于14小时/天,花粉能育。主要是由于长日照促进了IAA氧化酶的活性,分解IAA多而败育。 6、促进果实生长,诱导单性结实 雌蕊受精后,产生大量的IAA,吸收营养器官的养分运到子房,促进果实的生长。若在授粉前,用IAA喷洒柱头,可不经授粉而引起子房膨大,发育成无籽果实。用10mg/L的2,4-D喷洒番茄、茄子、辣椒、西瓜,可产生无籽果实。
7、抑制落花、落叶和落果。 一般认为当远轴端IAA含量大于近轴端时,叶片不脱落,反之则脱落。生产上用10mg/LNAA可使棉花保铃保蕾,洒施葡萄时,除产生无籽果实外,还可提高座果率。 六、生长素促进伸长的作用机理 (一)酸促生长学说
结论: 酸性溶液能促进细胞的伸长,生长素可起到酸性pH同样的作用 。 进一步实验表明,生长素促进细胞的伸长与细胞的代谢有关。两者的区别: 1)IAA诱导细胞壁塑性的增加只对活细胞有效,而H+所诱导的这种效应对死、活细胞都有效。 2)IAA诱导细胞的伸长是一个需能过程,CN-、二硝基酚可抑制生长素的这种效应,但对H+诱导的伸长则无影响。
实验2: 若将燕麦胚芽鞘横放,在其尖端挂上一个重量,使之伸长。当移去重量时,胚芽鞘只能部分恢复原状。我们将可逆的伸展能力称弹性。不可逆的伸展能力称塑性。试验指出生长素引起生长的增加幅度与细胞壁塑性变化相接近,生长素主要引起细胞塑性的增加。 胚芽鞘 F 原长 塑性 弹性
酸促生长学说(1970年由Rayle and Cleland提出) 认为: (1) 生长素和质膜 上的受体结合,活化了质子泵,将H+从细胞质 分泌到细胞壁,使细胞壁酸化。 (2)酸化的细胞壁一方面可使壁中酸不稳定的氢键易断裂,如木葡聚糖聚合体和纤维素微纤丝之间的氢键,另一方面可激活细胞壁中的一些水解酶的活性,如水解果胶质的β-半乳糖苷酶活性在pH4-5时比pH7时高3-10倍,β-1,4-葡聚糖酶的活性用IAA处理后,活性提高100倍。 (3)细胞壁多糖分子间结构交织点被破坏,联系松驰,可塑性增加。
由于生长素与H+-ATPase结合和随之带来的H+的主动分泌都需要一定的时间,所以IAA所引起伸长有10-15 min的滞后期,比酸所引起伸长的滞后期1min长。 (二)基因活化学说: 许多实验证明,生长素诱导的细胞伸长还与它能促进新原生质和细胞壁物质的合成有关,这种过程能持续几个小时。 核酸合成抑制剂放线菌素-D和蛋白质合成抑制剂亚胺环已酮可抑制IAA对伸长生长的诱导作用。
基因活化学说认为: 1、IAA与质膜上的受体结合后’诱发肌醇三磷酸IP3的产生。 2、IP3打开细胞器的Ca2+-Channel,释放内质网和液胞的Ca2+,这样就增加了细胞溶质中的Ca2+浓度。 3、细胞质中Ca2+浓度的增加刺激了蛋白激酶的活性,使蛋白质磷酸化。 4、活化的蛋白质因子与生长素结合,形成蛋白质-生长素复合物,转达移到细胞核,再活化特殊的mRNA,最后在核糖体上合成新的蛋白质。 5、另外Ca2+进入液胞,置换出H+,刺激H+-ATPase,向细胞壁分泌H+,增加细胞壁的塑性。
(三)IAA受体 1、IAA受体:是指能特异地识别IAA并能与其高度结合,进一步引起一系列生理生化变化的物质,称IAA受体。 2、 IAA受体定位 1)质膜上:如玉米胚芽中分离出了一种质膜IAA结合蛋白,Mr:40000,含两个亚基。质膜生长素结合蛋白(ABP)可促进质膜H+-ATPase的活性,从而促进伸长生长。 2)存在于细胞质中的内质网、液胞膜或核膜上: 在烟草愈伤组织中鉴定出了一种存在于泡液和细胞核内的ABP,这些ABP能显著促进生长素诱导的mRNA的增加。 最近在内质网上分离出一种ABP1,是分子量为22000糖蛋白,其与IAA相互作用可使原生质膨胀以及H+泵活化,而认为ABP1是IAA受体。
第二节 赤霉素(Gibberellins) 一、赤霉素的发现: GA是日本人黑泽英一,在1926年从水稻恶苗病的研究发现的。患恶苗病的水稻植物会发生徒长,是由赤霉菌的分泌物引起的。 1938年日本人薮田贞次郎等从水稻赤霉菌中分离到了赤霉素结晶。不久该项研究被二战中断,战后日本的赤霉素研究介绍到国外,立即引起了各国的重视。1959年确定了其化学结构。 GA3是第一个被分离到的赤霉素。目前证明,GA广泛存在于高等植物中,现在已知有125种GA。在真菌、藻类、蕨类、裸子、种子植物中都有存在赤霉素。
二、赤霉素的结构、体内分布和运输 (一)赤霉素的结构 GA是由4个异戍二烯基单位构成的双萜类物质。有一个基本结构—赤霉烷环。赤霉烷环上双键和-OH数目和位置的不同以及有无内酯环,就形成了不同的赤霉素。 依GA分子中碳原子总数的不同,植物体内有2类:C20-GA和C19-GA,C19-GA的活性大于C20-GA。公认植物体内最有活性的是GA1。GA3由赤霉菌分泌而来,可大量生产。理论研究常用GA1。 GA活性与分子结构有关:若有活性的,C3和C13位上有-OH,若失活,则C2上-OH.
(二)体内分布 广泛存在于植物组织。一般生长旺盛的组织器官如根尖、茎尖、幼果和正在发育的种子含量较高;生殖器官含量高于营养器官。量值为1-1000 ng/gFW。 (三)体内运输: GA在植物体内的运输没有极性。根尖合成的GA可经木质部向上运输,而叶原基产生的GA则可通过韧皮部向下运输,运输速度与光合产物相似为50-100 cm/h。但不同植物差异较大,如矮生豌豆为5 cm/h;马铃署为0.42mm/h。 三、GA的生物合成和代谢 (一)GA的代谢 GA合成后在植物体内的降解很慢,容易转变成无活性的束缚型GA。常见的束缚型GA有:GA-葡萄糖酯和GA-葡萄糖苷等。
束缚型GA是GA的贮藏和运输形式。在植物的不同发育时期,游离型GA和束缚型GA可以相互转化。如在种子成熟时,游离型GA不断转变成束缚型GA而贮藏起来,而在种子萌发期,束缚型GA通过酶解不断形成游离型GA调节生理代谢。束缚型GA是GA的贮藏和运输形式。在植物的不同发育时期,游离型GA和束缚型GA可以相互转化。如在种子成熟时,游离型GA不断转变成束缚型GA而贮藏起来,而在种子萌发期,束缚型GA通过酶解不断形成游离型GA调节生理代谢。 (二)生物合成场所 植物体内合成GA的场所是顶端幼嫩部分:根尖、茎尖、幼果和生长中的种子等。是在细胞的质体中如叶绿体内合成的。 最新的研究方法:在拟南芥中发现了一个可被GA活化的启动子。该启动子可使报告基因GUS (β-葡萄糖苷酸酶基因)表达,高等植物无此基因,根据报告基因GUS就可确定GA的合成部位。已知,花粉粒、根尖、茎尖、侧根原基、生长中的种子和果实都能合成GA。
(二)合成途径: 同位素示踪实验证明,GA是由乙酰COA经萜类生物合成过程,形成甲羟戊酸(MVA),再经牝牛儿焦磷酸、法呢基焦磷酸等,形成内根贝壳杉烯,由内根贝壳杉烯再经过系列转变形成GA12-7醛。再由GA12-7醛转化成各种不同的GA。GA12-7醛是GA的共同前体。可分三个阶段: (1)环化过程,形成贝壳杉烯 : 内根贝壳杉烯合成酶A 内根贝壳杉烯合成酶B MVA CPP 内根贝壳杉烯 [福斯方-D、AMO-1618、CCC、Pix] [Pix 、CCC] CH2OPP MVA 异烯基焦磷酸
(2)C-19的氧化 贝壳杉烯加氧酶 内根贝壳杉烯 根贝壳杉烯酸 GA12-7醛 。 在贝壳杉烯加氧酶催化下,C-19的-CH3经-CH2OH、-CHO、 -COOH的转变形成内根贝壳杉烯酸。 贝壳杉烯加氧酶:需O2和NADPH2和CytP450为辅助因子。PP333(多效唑或氯丁唑 )、B9等物可抑制CytP450的作用,而抑制GA的合成。
(3)GA12-7醛 转化为其它GA 1)GA12-7醛 C20-GA (GA12)。GA12-7醛的7-CHO 7-COOH, 就转变为GA12。 2)GA12-7醛 C19-GA:有2条途径: I) 13β-OH,产生有活性的GA: 13β-OH O2氧化 脱羧CO2 3β-OH GA12-7醛 GA53 GA19 GA20GA1 (C20) (C19)
若 2β-OH,GA1 GA8(无活性) II)不经 13β-OH,产生无活性的GA。例GA51。 GA的生物合成受CCC、B9、缩节胺Pix、多胶唑 PP333、福斯方和AMO1618的抑制。这些物质由于抑制了GA的合成,茎部近顶端分生组织细胞伸长受抑,使节间缩短,株形矮小。此类物质称生长延缓剂。
四、GA的生理效应 (一)促进茎的伸长 GA也能促进植物的伸长生长。且具下列特点: 1)能促进整株的植物生长,对矮生植物效果特别明显。原因是矮生品种基因发生了突变,GA生物合成受阻。但对离体茎段伸长的促进作用不明显,这一点与IAA不同。 2)能促进节间的伸长,不增加节数。 3)GA浓度很高,仍表现出最大的促进效应,不存在超最适浓度的抑制作用。 4)GA对根的伸长无促进作用。 拟南芥和水稻都有GA缺失突变体,植物矮化。外源施GA,植株可恢复到正常高度。育种时只要控制GA的合成就可得到矮生品种。水稻育秧用CCC和AMO-1618等可壮秧。百合花过高观赏性差,用多效唑处理,可增加观赏性。