Download

1 / 74
740 likes | 1.01k Views
PROTEIN SYNTHESIS. Protein. Protein tersusun atas satuan yang berupa asam amino . Jumlah asam amino yang umum terdapat pada jasad hidup ada 20 macam. Protein.

E N D
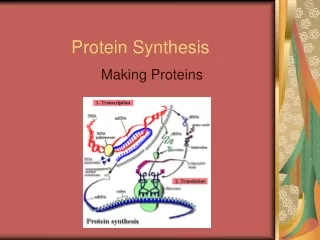
PROTEIN SYNTHESIS KRT-2011
Protein Protein tersusun atas satuan yang berupa asam amino. Jumlah asam amino yang umum terdapat pada jasad hidup ada 20 macam. KRT-2011
Protein Satu asam amino terdiri atas satu gugus amino, satu gugus karboksil, satu atom hidrogen, dan satu rantai samping yang terikat pada atom karbon. Susunan tetrahedral keempat gugus tersebut menentukan aktivitas optik asam amino sehingga ada dua bentuk isomer yaitu L-isomer dan D-isomer. KRT-2011
Struktur protein Dapat dibedakan dalam empat aras (level). • Struktur primermenyatakan susunan linear asam-asam amino sepanjang rantai polipeptida. • Struktur sekunder menggambarkan pola pelipatan (folding) bagian-bagian polipeptida ke dalam struktur yang teratur, misalnya heliks dan lembaran terlipat-β (β pleated sheet). • Struktur tersier menggambarkan pelipatan bagian-bagian antara heliks-α dan lembaran- β serta semua interaksi nonkovalen yang menyebabkan terjadinya pelipatan yang sesuai pada suatu rantai polipeptida. lnteraksi nonkovalen tersebut antara lain ikatan hidrogen, ikatan hidrofobik, dan interaksi van der Waals. • Struktur kuaterner, menunjukkan interaksi nonkovalen yang mengikat beberapa rantai polipeptida ke dalam satu molekul tunggal protein, misalnya hemoglobin. KRT-2011
Protein • Molekul yg sangat vital untuk organisme terdapt di semua sel • Polimer disusun oleh 20 mcm asam amino standar • Rantai asam amino dihubungkan dg iktn kovalen yg spesifik • Struktur & fungsi ditentukan oleh kombinasi, jumlah dan urutan asam amino • Sifat fisik dan kimiawi dipengaruhi oleh asam amino penyusunnya KRT-2011
Fungsi Protein • Reaksi kimia enzymes • Immune system antibodies • Mechanical structure tendons • Generation of force muscles • Nerve conduction ion channels • Vision eye lens • . . . and much more! KRT-2011
Fungsi Protein • Sebagai enzim Hampir semua reaksi biologis dipercepat atau dibantu oleh suatu senyawa makromolekul spesifik yang disebut enzim, dari reaksi yang sangat sederhana seperti reaksi transportasi karbon dioksida sampai yang sangat rumit seperti replikasi kromosom. Protein besar peranannya terhadap perubahan-perubahan kimia dalam sistem biologis. KRT-2011
Alat pengangkut dan penyimpan Banyak molekul dengan MB kecil serta beberapa ion dapat diangkut atau dipindahkan oleh protein-protein tertentu. Misalnya hemoglobin mengangkut oksigen dalam eritrosit, sedangkan mioglobin mengangkut oksigen dalam otot. • Pengatur pergerakan Protein merupakan komponen utama daging, gerakan otot terjadi karena adanya dua molekul protein yang saling bergeseran. KRT-2011
Penunjang mekanis Kekuatan dan daya tahan robek kulit dan tulang disebabkan adanya kolagen, suatu protein berbentuk bulat panjang dan mudah membentuk serabut. • Pertahanan tubuh atau imunisasi Pertahanan tubuh biasanya dalam bentuk antibodi, yaitu suatu protein khusus yang dapat mengenal dan menempel atau mengikat benda-benda asing yang masuk ke dalam tubuh seperti virus, bakteri, dan sel-sel asing lain. KRT-2011
Media perambatan impuls syaraf Protein yang mempunyai fungsi ini biasanya berbentuk reseptor, misalnya rodopsin, suatu protein yang bertindak sebagai reseptor penerima warna atau cahaya pada sel-sel mata. • Pengendalian pertumbuhan Protein ini bekerja sebagai reseptor (dalam bakteri) yang dapat mempengaruhi fungsi bagian-bagian DNA yang mengatur sifat dan karakter bahan KRT-2011
Sifat-sifat fisikokimia protein • Sifat fisikokimia setiap protein tidak sama, tergantung pada jumlah dan jenis asam aminonya. • Berat molekul protein sangat besar • Ada protein yang larut dalam air, ada pula yang tidak dapat larut dalam air, tetapi semua protein tidak larut dalam pelarut lemak. KRT-2011
Bila dalam suatu larutan protein ditambahkan garam, daya larut protein akan berkurang, akibatnya protein akan terisah sebagai endapan. Peristiwa pemisahan protein ini disebut salting out. • Apabila protein dipanaskan atau ditambahkan alkohol maka protein akan menggumpal. • Protein dapat bereaksi dengan asam dan basa KRT-2011
Initiation of protein synthesis Each ribosome has three binding sites for tRNAs; an A site where the incoming aminoacyl-tRNA binds, a P site where the tRNA linked to the growing polypeptide chain is bound, and an E site which binds tRNA prior to its release from the ribosome. Translation in prokaryotes begins by the formation of a 30S initiation complex between the 30S ribosomal subunit, mRNA, initiation factors and fMet tRNAf Met The 30S subunit binds to the Shine–Dalgarno sequence which lies 5’ to the AUG Start codon and is complementary to the 16S rRNA of the small ribosomal subunit. The ribosome then moves in a 3’ direction along the mRNA until it encounters the AUG codon. The 50S ribosomal subunit now binds to the 30S initiation complex to form the 70S initiation complex. In this complex, the anticodon of the fMet tRNAf Met is base paired to the AUG initiation codon (start codon) in the P site. KRT-2011
Elongation The elongation cycle consists of three steps: aminoacyl-tRNA binding, peptide bond formation, and translocation. In the first step, the aminoacyl-tRNA corresponding to the second codon binds to the A site on the ribosome as an aminoacyl-tRNA/EF-Tu/GTP complex. After binding, the GTP is hydrolyzed and EF-Tu/GDP is released. The EF-Tu is regenerated via the EF-Tu–EF-Ts exchange cycle. Peptide bond formation is catalyzed by peptidyl transferase between the C-terminus of the amino acyl moiety in the P site and the amino group of the aminoacyl-tRNA in the A site. In the final (translocation) step, EF-G/GTP binds to the ribosome, the deacylated tRNA moves from the P site to the E site, the dipeptidyl-tRNA in the A site moves to the P site, and the ribosome moves along the mRNA to place the next codon in the A site. The GTP is hydrolyzed to GDP and inorganic phosphate. When the next aminoacyl-tRNA binds to the A site in the next round of elongation, the deacylated tRNA is released from the E site KRT-2011
Termination The appearance of a UAA or UAG termination (stop) codon in the A site causes release factor RF1 to bind whereas RF2 recognizes UGA. RF3 assists RF1 and RF2. The release factors trigger peptidyl transferase to transfer the polypeptide to a water molecule instead of to aminoacyl-tRNA. The polypeptide, mRNA, and free tRNA leave the ribosome and the ribosome dissociates into its subunits ready to begin a new round of translation KRT-2011
PROTEIN TARGETING Both in prokaryotes and eukaryotes, newly synthesized proteins must be delivered to a specific subcellular location or exported from the cell for correct activity. This phenomenon is called protein targeting. KRT-2011
Secretory proteins Secretory proteins have an N-terminal signal peptide which targets the protein to be synthesized on the rough endoplasmic reticulum (RER). During synthesis it is translocated through the RER membrane into the lumen. Vesicles then bud off from the RER and carry the protein to the Golgi complex, where it becomes glycosylated. Other vesicles then carry it to the plasma membrane. Fusion of these transport vesicles with the plasma membrane then releases the protein to the cell exterior. KRT-2011
Plasma membrane proteins Plasma membrane proteins are also synthesized on the RER but become inserted into the RER membrane (and hence ultimately the plasma membrane) rather than being released into the RER lumen. The plasma membrane protein may pass once through the plasma membrane (Type I and Type II integral membrane proteins) or may loop back and forth, passing through many times (Type III integral membrane protein). The orientation of the protein in the membrane is determined by topogenic sequences within the polypeptide chain. KRT-2011
Type I proteins have a cleaved N-terminal signal sequence and a hydrophobic stop-transfer sequence, Type II have an uncleaved N-terminal signal sequence that doubles as the membrane-anchoring sequence, and Type III have multiple signal sequences and stop-transfer sequences. Proteins destined to be anchored in the membrane by a glycosyl- phosphatidylinositol (GPI) structure have both a cleaved N-terminal signal sequence and a C-terminal hydrophobic sequence that directs addition of the preformed GPI anchor. KRT-2011
Proteins of the endoplasmic reticulum Proteins destined for the RER have an N-terminal signal peptide, are synthesized on the RER, are translocated into the RER lumen or inserted into the RER membrane. C-terminal amino acid sequences (KDEL in soluble RER lumen proteins, KKXX in type I integral membrane proteins) are recognized by specific receptor proteins and retain the proteins in the ER KRT-2011
Lysosomal proteins Lysosomal proteins are targeted to the lysosomes via the addition of a mannose 6-phosphate signal that is added in the cis-compartment of the Golgi and is recognized by a receptor protein in the trans-compartment of the Golgi. The protein is then transported by specialized vesicles to a late endosome that later matures into a lysosome. The mannose 6-phosphate receptor recycles back to the Golgi for re-use. KRT-2011
Mitochondrial and chloroplast proteins Most mitochondria and chloroplast proteins are made on free cytosolic ribosomes, released into the cytosol and then taken up into the organelle. Uptake into the mitochondrial matrix requires a matrix-targeting sequence and occurs at sites where the outer and inner mitochondrial membranes come into contact. The process is mediated by hsp70 and hsp60 proteins and requires both ATP hydrolysis and an electrochemical gradient across the inner mitochondrial membrane. Targeting of proteins to other compartments of mitochondria or chloroplasts requires two signals. KRT-2011
Nuclear proteins Proteins destined for import into the nucleus typically require a nuclear localization signal, four to eight amino acids long, located internally in the protein. Uptake occurs via nuclear pores and requires ATP hydrolysis. KRT-2011
Review : Replication The DNA double helix acts as a template for its own duplication. Because the nucleotide A will pair successfully only with I and G only with C, each strand of DNA can serve as a template to specify the sequence of nucleotides in its complementary strand by DNA basepairing. In this way, a double-helical DNA molecule can be copied precise KRT-2011
Review : Replication The chemistry of DNA synthesis. The addition of a deoxyribonucleotide to the 3' end of a polynucleotide chain (the primer strand) is the fundamental reaction by which DNA is synthesized. As shown, base-pairing between an incoming deoxyribonucleoside triphosphate and an existing strand of DNA (the template strand) guides the formation of the new strand of DNA and causes it to have a complementary nucleotide sequence. KRT-2011
Review : Replication The semiconservative nature of DNA replication. In a round of replication, each of the two strands of DNA is used as a template for the formation of a complementary DNA strand. The original strands therefore remain intact through many cell generations KRT-2011
The structure of a DNA replication fork. Because both daughter DNA strands are polymerized in the 5’ to 3'direction, the DNA synthesized on the lagging strand must be made initially as a series of short DNA molecules, called Okazaki fragments. On the lagging strand, the Okazaki fragments are synthesized sequentially, with those nearest the fork being the most recently made. KRT-2011
The synthesis of one of many DNA fragments on the lagging strand. In eucaryotes, RNA primers are made at intervals spaced by about 200 nucleotides on the lagging strand, and each RNA primer is approximately 10 nucleotides long. This primer is erased by a special DNA repair enzyme (an RNAse H) that recognizes an RNA strand in an RNA/DNA helix and fragments it; this leaves gaps that are filled in by DNA polymerase and DNA liqase KRT-2011
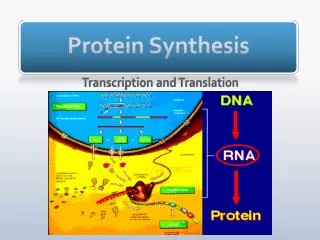
FROM DNA TO RNA The pathway from DNA to protein. The flow of genetic information from DNA to RNA (transcription) and from RNA to protein (translation) occurs in all living cells Transcription and translation are the means by which cells read out, or express, the genetic instructions in their genes. Because many identical RNA copies can be made from the same gene, and each RNA molecule can direct the synthesis of many identical protein molecules, cells can synthesize a large amount of protein rapidly when necessary. But each gene can also be transcribed and translated with a different efficiency, allowing the cell to make vast quantities of some proteins and tiny quantities of others KRT-2011
How Cells Read the Genome: From DNA to Protein Genes can be expressed with different efficiencies. In this example, gene A is transcribed and translated much more efficiently than gene B. This allows the amount of protein A in the cell to be much greater than that of protein B. KRT-2011
Portions of DNA Sequence Are Transcribed into RNA (transcription.) The first step a cell takes in reading out a needed part of its genetic instructions is to copy a particular portion of its DNA nucleotide sequence—a gene—into an RNA nucleotide sequence. The information in RNA, although copied into another chemical form, is still written in essentially the same language as it is in DNA—the language of a nucleotide sequence KRT-2011
Struktur RNA RNA can fold into specific structures. RNA is largely single-stranded, but it often contains short stretches of nucleotides that can form conventional base pairs with complementary sequences found elsewhere on the same molecule. These interactions, along with additional “nonconventional” base-pair interactions, allow an RNA molecule to fold into a three-dimensional structure that is determined by its sequence of nucleotides. <AATC> (A) Diagram of a folded RNA structure showing only conventional base-pair interactions. (B) Structure with both conventional (red) and nonconventional (green) base-pair interactions. (C) Structure of an actual RNA, a portion of a group I intron. Each conventional base-pair interaction is indicated by a “rung” in the double helix. Bases in other configurations are indicated by broken rungs. KRT-2011
Review : Transcription KRT-2011
Before the synthesis of a particular protein can begin, the corresponding mRNA molecule must be produced by transcription. Bacteria contain a single type of RNA polymerase (the enzyme that carries out the transcription of DNA into RNA). An mRNA molecule is produced when this enzyme initiates transcription at a promoter, synthesizes the RNA by chain elongation, stops transcription at a terminator, and releases both the DNA template and the completed mRNA molecule. In eucaryotic cells, the process of transcription is much more complex, and there are three RNA polymerases polymerase I, II, and III—that are related evolutionarily to one another and to the bacterial polymerase. KRT-2011
FROM RNA TO PROTEIN An mRNA Sequence Is Decoded in Sets of Three Nucleotides Once an mRNA has been produced by transcription and processing, the information present in its nucleotide sequence is used to synthesize a protein. Transcription is simple to understand as a means of information transfer: since DNA and RNA are chemically and structurally similar, the DNA can act as a direct template for the synthesis of RNA by complementary base-pairing. KRT-2011
Genetic code In contrast, the conversion of the information in RNA into protein represents a translation of the information into another language that uses quite different symbols. Moreover, since there are only 4 different nucleotides in mRNA and 20 different types of amino acids in a protein, this translation cannot be accounted for by a direct one-to-one correspondence between a nucleotide in RNA and an amino acid in protein. The nucleotide sequence of a gene, through the intermediary of mRNA, is translated into the amino acid sequence of a protein by rules that are known as the genetic code KRT-2011
Genetic code Any series of three bases (or nucleotides) in the DNAprescribes for an amino acid in the protein chain, or gives a ‘stop transcribing’ signal. The bases are always read from left to right. The chain usually starts with ATG or methionine (Met). Abbreviations used: A, adenine; G, guanine; C, cytosine; T, thymine (or U, uracil in RNA). Ala, alanine; Arg, arginine; Asn, asparagine; Asp, aspartic acid; Cys, cysteine; Gln, glutamine; Glu, glutamic acid; Gly, glycine; His, histidine; Ile, isoleucine; Leu, leucine; Lys, lysine; Met, methionine; Phe, phenylalanine; Pro, proline; Ser, serine; Thr, threonine; Trp, tryptophan; Tyr, tyrosine; Val, valine KRT-2011
Kode genetik Di alam ada 20 macam asam amino yang umum terdapat di dalam struktur polipeptida jasad hidup. Masing-masing asam amino mempunyai kodon yang spesifik sedangkan nukleotida hanya ada 4 macam yaitu A, U, G, dan C (Tabel1 2.2). Jika suatu kodon hanya terdiri atas dua nukleotida maka hanya akan ada 42 = 16 asam amino, tetapi apabila kodon disusun oleh 3 nukleotida maka akan diperoleh 43 (= 64) asam amino. S Sedangkan jumlah asam amino yang umum diketahui ada pada jasad hidup hanya 20 macam. Beberapa kodon diketahui mengkode asam amino yang sama. Fenomena ini dikenal sebagai genetic code redundancy (degeneracy) KRT-2011
Tranlasi Translasi adalah proses penerjemah urutan nucleotida yang ada pada molekul mRNA menjadi rangkaian asam-asam amino yang menyusun suatu polipeptida atau protein. Hanya molekul mRNA yang ditranslasi, sedangkan rRNA dan tRNA tidak ditranslasi. Molekul mRNA merupakan transkrip (salinan) urutan DNA yang menyusun suatu gen dalam bentuk ORF (open reading frame, kerangka baca terbuka) Molekul rRNA adalah salah satu molekul penyusun ribosom, yakni organel tempat berlangsungnya sintesis protein, tRNA adalah pembawa asam-asam amino yang akan disambungkan menjadi rantai polipeptida KRT-2011
Reading frames • Suatu ORF dicirikan oleh: (1) kodoninisiasi translasi, yaitu urutan ORF ATG (pada DNA) atau AUG (pada mRNA), (2) serangkaian urutan nukleotida yang menyusun banyak kodon, dan (3) kodon terminasi translasi, yaitu TAA (UAA pada mRNA). TAG (UAG pada mRNA) atau TGA (UGA pada mRNA) The three possible reading frames in protein synthesis. In the process of translating a nucleotide sequence (blue) into an amino acid sequence (red), the sequence of nucleotides in an mRNA molecule is read from the 5’ end to the 3’ end in consecutive sets of three nucleotides. In principle, therefore, the same RNA sequence can specify three completely different amino acid sequences, depending on the reading frame. In reality, however, only one of these reading frames contains the actual message. KRT-2011
Kodon (kode genetik) Kodon (kode genetik) adalah urutan nukleotida yangterdiri atas 3 nukleotida yanq berurutan (sehingga sering disebut sebagai triplet codon, yang menyandi suatu kodon asam amino tertentu, misalnya urutan ATG (AUG pada mRNA) mengkode asam amino metionin, Kodon inisiasi translasi merupakan kodon untuk asam amino metionin yang mengawali struktur suatu polipeptida (protein). Pada prokaryot, asam amino awal tidak berupa metionin tetapi formil metionin (fMet). Wobble base-pairing between codons and anticodons. If the nucleotide listed in the first column is present at the third, or wobble, position of the codon, it can base-pair with any of the nucleotides listed in the second column. Thus, for example, when inosine (I) is present in the wobble position of the tRNA anticodon, the tRNA can recognize any one of three different codons in bacteria and either of two codons in eucaryotes. The inosine in tRNAs is formed from the deamination of guanine (see Figure 6–55), a chemical modification that takes place after the tRNA has been synthesized KRT-2011
Dalam proses translasi, rangkaian nukleotida pada mRNA akan dibaca tiap tiga nukleotida sebagai satu kodon untuk satu asam amino, dan pembacaan dimulai dari urutan kodon metionin (ATG pada DNA atau AUG pada RNA) KRT-2011
Ada beberapa aspek yang perlu diketahui mengenai kode genetik, yaitu: Kode genetik bersifat tidak saling tumpang-tindih (non-overlappind kecuali pada kasus tertentu, misalnya pada bakteriofag Tidak ada sela (gap) di antara kodon satu dengan kodon yang lain. Tidak ada koma di antara kodon. Kodon bersifat degenerotea, buktinya ada beberapa asam amino yang mempunyai lebih dari satu kodon. Secara umum, kodon bersifat hampir universal karena pada beberapa organel jasad tinggi ada beberapa kodon yang berbeda dari kodon yang digunakan pada sitoplasm KRT-2011
Dalam proses translasi, setiap kodon berpasangan dengan antikodon yang sesuai yang terdapat pada molekul tRNA. Sebagai contoh, kodon metionin (AUG) mempunyai komplemennya dalam bentuk antikodon UAC yang terdapat pada tRNAMet Pada waktu tRNA yang membawa asam amino diikat ke dalam sisi A pada ribosom, maka bagian antikodonnya berpasangan dengan kodon yang sesuai yang ada pada sisi A tersebut. Oleh karena itu, suatu kodon akan menentukan asam amino yang disambungkan ke dalam polipeptida yang sedang disintesis di dalam ribosom KRT-2011
Translasi berlangsung di Ribosom Translasi berlangsung di dalam ribosom. Ribosom disusun oleh molekul rRNA dan beberapa macam protein. Ribosom tersusun atas dua yaitu subunit kecil dan subunit besar. The RNA-binding sites in the ribosome. Each ribosome has one binding site for mRNA and three binding sites for tRNA: the A-, P-, and E-sites (short for aminoacyl-tRNA, peptidyl-tRNA, and exit, respectively). KRT-2011
Transkripsi - Tranlasi Pada jasad prokaryot, translasi sudah dimulai sebelum proses transkripsi (sintesis mRNA) selesai dilakukan. Dengan demikian, proses transkripsi dan translasi pada prokaryot berlangsung secara hampir serentak. Sebaliknya pada eukoryot, proses translasi baru dapat berlangsung jika proses transkripsi (sintesis mRNA yang matang) sudah selesai dilakukan. Hal ini disebabkan oleh perbedaan dalam hal struktur sel antara prokaryot dengan eukaryot KRT-2011
Proses Translasi Proses translasi berlangsung melalui tiga tahapan utama, yaitu: (1 ) inisiasi (initiation), (2) pemanjangan (elongation) poli-asam amino, dan (3) pengakhiran (termination) translasi. Oleh karena itu, ada sekitar 20 macam tRNA yang masing-masing membawa asam amino spesifik, karena di alam ada sekitar 20 asam amino yang menyusun protein alami. Masing-masing asam amino diikatkan pada tRNA yang spesifik melalui proses yang disebut sebagaiI RNA charging (penambahan muatan berupa asam amino) KRT-2011
Aminoasil tRNA The structure of the aminoacyl-tRNA linkage. The carboxyl end of the amino acid forms an ester bond to ribose. Because the hydrolysis of this ester bond is associated with a large favorable change in free energy, an amino acid held in this way is said to be activated. (A) Schematic drawing of the structure. The amino acid is linked to the nucleotide at the 3’ end of the tRNA (B) Actual structure corresponding to the boxed region in (A). There are two major classes of synthetase enzymes: one links the amino acid directly to the 3¢-OH group of the ribose, and the other links it initially to the 2’-OH group. In the latter case, a subsequent transesterification reaction shifts the amino acid to the 3’ position. As in Figure 6–56, the “R group” indicates the side chain of the amino acid Sebelum inisiasi translasi di lakukan, diperlukan molekul tRNA (aminoasil tRNA) yang berfungsi membawa asam amino spesifik. KRT-2011
Inisiasi translasi (eukariyot) Kodon inisiasi adalah metionin Molekul tRNA inisiator disebut sebagai tRNAiMet. Ribosom bersama-sama dengan tRNAiMet dapat menemukan kodon awal dengan cara berikatan dengan ujung 5' (tudung), kemudian melakukan pelarikan (scanning) transkrip ke arah hilir (dengan arah 5' 3') sampai menemukankodon awal (AUG). Menurut model scanning tersebut, ribosom memulai translasi pada waktu menjumpai sekuens AUG yang pertama kali KRT-2011