Download
1 / 60
600 likes | 606 Views
This article explores various RNA-mediated gene silencing mechanisms, including homology-dependent gene silencing, transcriptional gene silencing, post-transcriptional gene silencing, and viral response in chordates. Additionally, it discusses the biogenesis of miRNAs and siRNAs, as well as the fate of mRNA loaded with the miRISC. The role of poly(A) and the cap in translation initiation, as well as the promotion of mRNA deadenylation by miRNAs, are also covered.

E N D
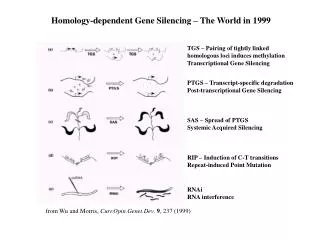
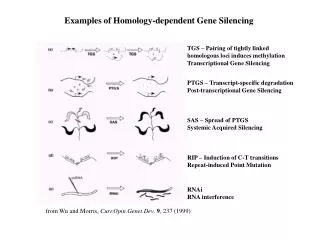
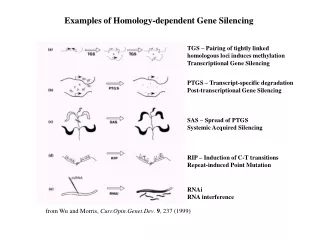
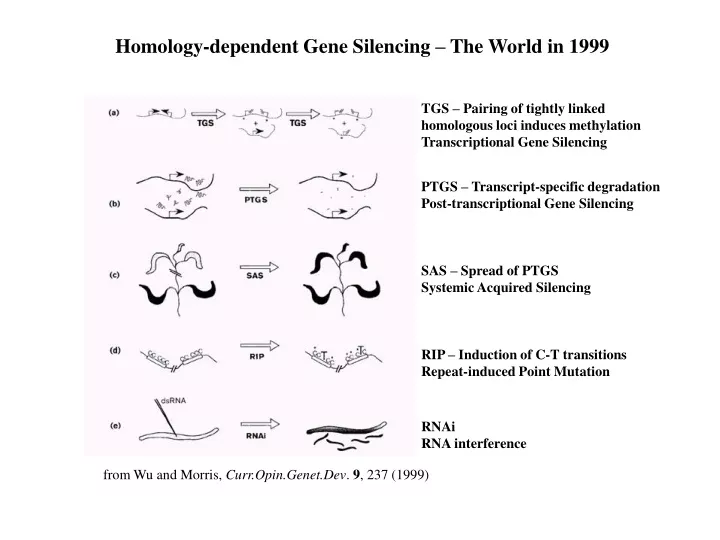
Homology-dependent Gene Silencing – The World in 1999 TGS – Pairing of tightly linked homologous loci induces methylation Transcriptional Gene Silencing PTGS – Transcript-specific degradation Post-transcriptional Gene Silencing SAS – Spread of PTGS Systemic Acquired Silencing RIP – Induction of C-T transitions Repeat-induced Point Mutation RNAi RNA interference from Wu and Morris, Curr.Opin.Genet.Dev. 9, 237 (1999)
Small RNAs from tenOever, Nature Rev.Microbiol. 11, 169 (2013)
Response to Virus Infection in Chordates Viral dsRNA is recognized by PRRs in the cytoplasm or TLRs in endosomes Induce expression of type I interferons Leads to transactivation of >250 genes Slows viral infection and allows time for an adaptive immune response from tenOever, Nature Rev.Microbiol. 11, 169 (2013)
viRNAs are an Antiviral Innate Immune System viRNAs are derived from the virus and loaded onto the RISC viRNAs bind the viral RNA target with perfect complementarity and eliminates the target Chordates do not produce viRNA from tenOever, Nature Rev.Microbiol. 11, 169 (2013)
Response of Mammalian Cells to Long dsRNA Long dsRNA induces interferon response in vertebrates PKR phosphorylates eIF2a to inhibit translation 2’-5-oligoadenylate synthase is induced, which activates RNaseL and leads to nonspecific mRNA degradation siRNA does not invoke the interferon response from McManus and Sharp, Nature Rev.Genet.3, 737 (2002)
The lin-14 Mutant has an Altered Pattern of Cell Division The PNDB neuroblast is generated prematurely The LIN-14 protein prevents L2-type cell divisions from Lodish et al., Molecular Cell Biology, 6th ed. Fig 21-6
miRNAs Regulate Development in C. elegans The LIN-14 protein prevents L2-type cell divisions During L2, lin-4 miRNA prevents translation of lin-14 mRNA In the adult, let-7 inhibits lin-14 and lin-41 translation Absence of LIN-41 permits lin-29 translation and generation of adult cell lineages from Lodish et al., Molecular Cell Biology, 6th ed. Fig 21-6
lin-4 Inhibits Translation of lin-14 mRNA Mutations in lin-4 disrupt regulation of larval development in C. elegans lin-4 antagonizes lin-14 function lin-4 encodes the precursor to a 22 nt- long microRNA that is partially complementary to sites in the 3’UTR of lin-14 mRNA Annealing of lin-4 to lin-14 mRNA inhibits translation from Li and Hannon, Nature Rev.Genet. 5, 522 (2004)
Biogenesis of miRNAs and siRNAs miRNAs are genomically encoded siRNAs are produced exogenously or from bidirectionally transcribed RNAs Drosha processes pri-miRNA to pre-miRNA in the nucleus miRNA is selectively incorporated into the RISC for target recognition Guide strand of siRNA is incorporated into the RISC for target recognition miRNAs have imperfect complementarity to their target mRNA and inhibit translation siRNAs form perfect duplex with their target mRNA and trigger mRNA degradation from Li and Hannon, Nature Rev.Genet. 5, 522 (2004)
Triggers of RNAi-Mediated Gene Silencing in Mammals from Mittal, Nature Rev.Genet. 5, 355 (2004)
Strand Selection Into the RISC The strand with its 5’-terminus at the less stable end of the duplex is incorporated into the RISC from Sontheimer, Nature Rev.Mol.Cell Biol.6, 127 (2005)
Strand Selection of Processed siRNA into the RISC The PAZ domain of Dicer binds to the pre-existing dsRNA end The strand that has its 3’-end bound to the PAZ domain preferentially assembles into the RISC from Sontheimer, Nature Rev.Mol.Cell Biol.6, 127 (2005)
Guide RNA Loading Onto Argonaute PAZ domain binds 3’-overhang 5’-end of guide RNA is anchored in a conserved pocket of the PIWI domain Argonaute slices passenger strand of siRNA from Parker and Barford, Trends Biochem.Sci. 31, 622 (2006)
Mechanisms of miRNA Sequence Diversification Seed shifting that results from variations in Drosha or Dicer processing generates isomiRs In arm shifting, mutations within the precursor change the ratio of miRNA to miRNA* loading In hairpin shifting, the folding is changed into a new configuration In cells containing adenosine deaminase, A is converted to I from Berezikov, Nature Rev.Genet. 12, 846 (2011)
The Fate of mRNA Loaded With the miRISC Targeted mRNA accumulates in P bodies mRNA is stored in P bodies, undergoes degradation, or reenters the translation pathway from Rana, Nature Rev.Mol.Cell Biol.8, 23 (2007)
Role of Poly(A) and Cap in Translation Initiation The cap structure is recognized by eIF4F Poly(A) is recognized by PABPC PABPC interacts with eIF4G Recruitment of the preinitiation complex is increased from Huntzinger and Izaurralde, Nature Rev.Genet. 12, 99 (2011)
miRNAs Promote mRNA Deadenylation miRNA guide strand associates with AGO AGO interacts with GW182 GW182 may compete with eIF4G for binding to PABPC and prevents mRNA circularization GW182 may reduce the affinity of PABPC for the poly(A) tail Assembly of AGO-GW182-PABPC complex triggers deadenylation by CAF1-CCR4-NOT from Huntzinger and Izaurralde, Nature Rev.Genet. 12, 99 (2011)
Fate of Deadenylated mRNAs Deadenylated mRNAs are stored in a translationally repressed state Deadenylated mRNAs are decapped by DCP2 associated with decapping activators Decapped mRNA is degraded by XRN1 from Huntzinger and Izaurralde, Nature Rev.Genet. 12, 99 (2011)
Overview of RNA-Mediated Gene Silencing siRNA siRNA triggers endonucleolytic cleavage of perfectly-matched complementary targets Cleavage is catalyzed by Argonaute proteins The resulting mRNA fragments are degraded miRNA miRNA triggers accelerated deadenylation and decapping of partially-complementary targets and requires Argonaute proteins and a P-body component miRNA represses translation from Eulalio et al., Nature Rev.Mol.Cell Biol.8, 9 (2007)
Secretion of miRNAs Specific miRNAs can be preferentially sorted into vesicles and delivered to recipient cells from Chen et al., Trends Cell Biol. 22, 125 (2012)
Regulation of siRNA Levels in C. elegans RNA-dependent RNA polymerase amplifies siRNA RRF-3 prevents siRNA amplification ERI-1 is an siRNA-specific RNase from Timmons, BioEssays26, 715 (2004)
Prevalence of and Regulation by miRNAs At least 1400 miRNA-encoding genes in humans miRNAs regulate ~50% of the human transcriptome miRNAs fine tune the expression of proteins in a cell
Organismal Complexity May Be Due to Differences in Regulation of Gene Expression Number of protein-coding genes are similar in animals There is a continuous acquisition of novel miRNAs during evolution Lineage-specific loss of miRNAs also occurs miRNA complexity correlates with an increase in morphological complexity There are now estimated to be 1,424 miRNAs in humans from Technau, Nature455, 1184 (2008)
let-7 is a Heterochronic Gene in C. elegans Mutations in heterochronic genes cause temporal cell fate transformations that are altered relative to the timing of events in other cells or tissues let-7 mutations cause an overproliferation of seam cells Overproliferation of cells is a characteristic of stem cells and cancer from Büssing et al., Trends Mol.Med. 14, 400 (2008)
Regulation of Differentiation by let-7 let-7 levels are reduced in stem cells Lin28 promotes reprogramming by inhibition of let-7 maturation from Viswanathan and Daley, Cell140, 445 (2010)
Reprogramming to iPS Cells Oct4 Sox2 Klf4 c-Myc Oct4 Sox2 NANOG Lin28 or Lin28 represses let-7 Is let-7 repression important for establishment of pleuripotent state? c-Myc is a let-7 target, so Lin28 replaces c-Myc Transfection of ESCC (ES cell-specific cell cycle-regulating) miRNAs can generate ES cells without protein-encoding factors
Links of let-7/Lin28 to Cancer let-7 is a tumor suppressor The oncogenes c-Myc, K-Ras, and cyclin D1 are let-7 targets Lin28 is an oncogene that is activated in 15% of human tumors Lin28 is also a let-7 target let-7 Lin28 double-negative feedback loop
Lin28 Prevents let-7 Maturation let-7 promotes differentiation Lin28a and Lin28b repress let-7 biogenesis by two distinct mechanisms Lin28a recruits TUTase which uridylates the miRNA and promotes let-7 degradation Lin28b inhibits Drosha- mediated processing of let-7 During differentiation, let-7 targets Lin28 mRNA, which reinforces developmental commitment from Thornton and Gregory, Trends Cell Biol. 22, 474 (2012)
Summary of Lin28 let-7 Regulation of Differentiation and Oncogenesis from Thornton and Gregory, Trends Cell Biol. 22, 474 (2012) Lin28 prevents let-7 muturation let-7 promotes differentiation and prevents transformation Lin28 promotes reprogramming or transformation ESCC miRNAs maintain Lin28 expression
A MicroRNA Regulates Neuronal Differentiation by Controlling Alternative Splicing miR-124 targets a component of a repressor of neuron-specific genes miR-124 results in reduced expression of PTBP1 leading to the accumulation of PTBP2 PTBP2 results in a global switch to neuron- specific alternative splicing patterns from Makeyev et al., Mol.Cell27, 435 (2007)
The Role of miRNA in Cancer miRNA profiles define the cancer type better than mRNA expression data miRNA expression is lower in cancers than in most normal tissues, but expression of some miRNAs is increased Down-regulation of all miRNAs enhanced tumor growth The undifferentiated state of malignant cells is correlated with a decrease in miRNA expression c13orf25 miRNA is the first non-coding oncogene, is upregulated by c-Myc, and is involved in leukemia development c13orf25 inhibits expression of E2F1, a cell cycle regulator from He et al., Nature435, 828 (2005) Lu et al., Nature435, 834 (2005) Lujambio and Lowe, Nature482, 347 (2012)
miRNAs and Breast Cancer Metastasis Loss of miR-126 and miR-355 when human breast cancer cells develop metastatic potential Restoring expression of these miRNAs in malignant cells suppresses metastasis in vivo miR-355 targets the progenitor cell transcription factor SOX4, and the ECM component tenascin C miR-10b and miR-9 induce metastasis from Tavasoie et al., Nature451, 147 (2008)
Role of MicroRNAs and Epigenetics in Cancer EZH2 (a PcG protein) overexpression promotes cell proliferation Expression of EZH2 is inhibited by miR-101 miR-101 expression decreases during prostate cancer progression from Varambally et al., Science 322, 1695 (2008) miR-29 inhibits DNMT3A and DNMT3B in lung cancer from Lujambio and Lowe, Nature482, 347 (2012)
Inhibition of Endogenous miRNA function miRNA sponges Vectors express multiple copies of miRNA target sites Endogenous miRNA is saturated and prevented from silencing its natural product Pseudogene transcripts can act as miRNA sponges from Brown and Naldini, Nature Rev.Genet. 10, 578 (2009)
Competitive Endogenous RNAs (ceRNAs) 70-90% of the human genome is transcribed, but less than 2% of the genome encodes protein-coding genes The human transcriptome contains 21.000 protein-coding genes, 9,000 small RNAs, 10,000-32,000 lncRNAs and 11,000 pseudogenes All RNA transcripts that contain miRNA binding sites that regulate each other by competing for shared miRNAs ceRNAs can fine-tune gene expression
Regulation of PTEN Levels by a Pseuodogene The expression level of PTEN is crucial for its tumor suppressive function PTEN expression is downregulated by miRNAs PTENP1 is a pseudogene which contains the same MRE in the 3’-UTR PTENP1 RNA is a ceRNA that enhances PTEN expression by competing for a shared miRNA from Rigoutsos, Nature465, 1016 (2010)
The PTEN ceRNA Network PTEN expression levels are regulated by a large network of miRNAs, mRNAs, and ceRNAs The PTEN ceRNA interactions are part of a regulatory layer comprising of more than 248,000 miRNA-mediated interactions from Tay et al., Nature505, 344 (2014)
Circular RNAs can be microRNA Sponges Human fibroblasts have 25,000 circRNAs derived from 15% of transcribed genes The splicing machinery is involved in circRNA biogenesis circRNAs are resistant to degradation triggered by miRNAs from Wilusz and Sharp, Science340, 440 (2013)
Immunostimulatory Effects of dsRNA Long dsRNA induces PKR Toll-like receptors in endosomes recognize dsRNA and activate the interferon response Blunt-ended dsRNA are recognized by RIG-1 helicase and activates the immune response from Kim and Rossi, Nature Rev.Genet. 8, 173 (2007)
DNA Vector-based RNAi from Shi, Trends Genet.19, 9 (2003)
The Design of Optimal siRNAs 21 nt RNA that contains 2 nt 3’- overhangs and phosphorylated 5’-ends Lower stability at the 5’-end of the antisense terminus Low stability in the RISC cleavage site Low secondary structure in the targeted region of the mRNA from Mittal, Nature Rev.Genet. 5, 355 (2004)
Delivery of siRNA for Therapy siRNA is not taken up by most mammalian cells Cholesterol-conjugated siRNA is taken up by the LDL receptor siRNA bound to targeted antibody linked to protamine can achieve cell-specific siRNA delivery from Dykxhoorn and Lieberman, Cell126, 231 (2006)
Cell-Specific Delivery of siRNA Fuse Fab targeting antibody with protamine siRNA binds noncovalently with protamine Complex is endocytosed into cells expressing the epitope siRNA is released from the endosome and enters the RISC from Rossi et al., Nature Biotechnol. 23, 682 (2005)
RNAi-dependent Chromatin Silencing in S. pombe Overlapping RNAs from centromeric region is processed into siRNA siRNA activates or recruits Clr3 methyltransferase that methylates H3 on K9 Deletion of RNAi pathway genes cause loss of silencing at centromeres and reduced H3 K9 methylation at centromeric regions from Allshire, Science297, 1818 (2002)
Small RNAs Modulate Viral Infection Viral-encoded miRNA facilitate viral infection and persistence Host cell-encoded miRNAs inhibit or facilitate viral replication Viral suppressors of RNA silencing (VSR) inhibit the RNAi pathway
Function of SV40 miRNA SV40 miRNA is synthesized late in the viral life cycle and targets TAg mRNA SV40 miRNA aids immune invasion by reducing susceptibility to lysis by CTLs Polyomaviruses also have viral miRNA that targets TAg Infection with Py mutant lacking the miRNA resulted in no difference in viral load or immune response from Sarnow et al., Nature Rev.Microbiol. 4, 651 (2006)
Effects of Adenovirus VA1 MicroRNA VA1 binds to and prevents PKR activation to inhibit the innate immune response VA1 competes with exportin-5 and inhibits Dicer to inhibit the RNAi pathway from Sarnow et al., Nature Rev.Microbiol. 4, 651 (2006)
A MicroRNA was Thought to Protect HSV-1-infected Neurons from Apoptosis LAT is the only viral gene expressed during latent infection in neurons miR-LAT is generated from the LAT gene miR-LAT downregulates TGF-b and SMAD3 and contributes to the persistence of HSV-1 in neurons in a latent form Paper retracted – 2008. Repeatedly unable to detect miRNA from Gupta et al., Nature 442, 82 (2006)
Cellular miRNAs Modulates Viral Infection PFV-1 replication is stimulated by a plant VSR implicating the role of small RNAs in the viral life cycle miR-32 inhibits viral replication Tas is a PFV-1-encoded protein that inhibits RNAi miR-122 increases HCV replication in the liver from Sarnow et al., Nature Rev.Microbiol. 4, 651 (2006) miR-122 stabilizes the HCV genome by binding the 5’-UTR
miR-122 Protects the HCV Genome From Degradation Xrn1 is a cytoplasmic exonuclease that normally degrades HCV RNA miR-122 increases HCV RNA stability by shielding the genome against Xrn1 miR-122 also enhances HCV RNA replication that is independent on its action against Xrn1 from Garcia-Sastre and Evans, Proc.Nat.Acad.Sci.110, 1571 (2013)