Download

1 / 68
700 likes | 961 Views
Gene Regulation during Development. Cells from different parts establish different program of gene expression , and most differential gene expression is regulated at the level of transcription initiation. This chapter ,we are going to learn gene Regulation during Development. Outline

E N D
Cells from different parts establish different program of gene expression , and most differential gene expression is regulated at the level of transcription initiation .
This chapter ,we are going to learn gene Regulation during Development Outline Section 1.Three strategies by which cells are instructed to express specific sets of gene during development Section 2. Example of the three strategies for establishing different gene expression Section 3. The molecular biology of Drosophila embryogenesis
Section 1 The three major ways strategies used to instruct genetically-identical cells to express distinct sets of genes and thereby differentiate into diverse cells types are : 1.mRNA localization 2.cell-to-cell contact 3.Signaling through the diffusion of a secreted signaling molecule
mRNA localization Principal: distributing mRNA asymmetrically during cell division→ daughter cells inheriting different amount of the regulator →following different pathways of development The roles of those mRNA are : encoding RNA-binding proteins or cell signaling molecules and transcriptional activators and repressors .
The common mechanism for localizing mRNAs is:an mRNA can be transported form one end to the other by means of an adapter protein ,which binds to a specific sequence within the 3’UTR of an mRNA and cytoskeleton. So the mRNA adapter complex can be moved with the cytoskeleton.
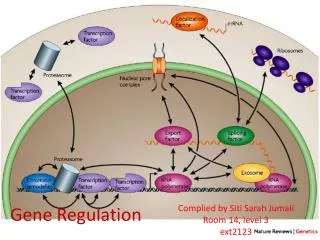
Cell to cell contact Principal: a cell producing extracellular signaling proteins, then the given signal is recognized by specific receptor on the surface of recipient cells and triggers changes in gene expression in the recipient.
This communication from the cell surface to the nucleus often involves signal transduction pathways . There are a few basic features of these pathways: 1. sometimes ligand-receptor interactions induce an enzymatic cascade the ultimately regulatory proteins already present in the nucleus.
2.activated receptor cause the release of DNA-binding proteins from the cell surface or cytoplasm into the nucleus
3.Upon cleavage,the intracytoplasmic domain of the receptor is released from the cell surface and enters the nucleus, to associate with DNA-binding proteins and then influence transcript
Signaling through the diffusion of a secreted signaling molecule
A recurring theme in development is the importance of a cell’s position within a developing embryo or organ in determining what it will become.The influence of location on development is called positional information.Signaling molecules that control position information are sometimes called morphogens
The most common ways of establishing positional information :A small group of cells synthesize andsecrete signals → the signal distribute in an extracellular gradient → cells in different positions receive different signal concentrations → levels of activated transcription factor differ → lead to the expression of different sets of genes
Section 2Examples of the Three Strategies for Establishing Differential Gene Expression
EXAMPLE 1 the localized Ash 1 repressor controls mating types yeast by silencing the HO gene
The yeast S.cerevisiae can grow as haploid cells that divide by budding. Which give rises to two cells:mother cell and the daughter cell.
SWITCHING ISCONTROLLED BY THE PRODUCT OF THE HOGENE A mother cell and its daughter cell can exhibit different mating types. This difference arises by a process called mating-type switching. After budding, a mother cell can switch mating type.
It is kept silent in the daughter cell due to the selective expression of a repressor called Ash1. The ash1 gene is transcribed in the mother cell during budding.But the encoded RNA becomes localized within the daughter cell through the following process.
During budding ,the ash1 mRNA attaches to the growing ends of microtubules .movement is directed and begins at the “-” ends of the filament and extends with the growing “+” ends.
The ash1 mRNA transport depends on the binding of the She2 and She3 adapter proteins to specific sequences contained within the 3’UTR
Once localized within the daughter cell, the ash1 mRNA is translated into a repressor protein that binds to, and inhibits the transcription of, the HO gene . This silencing of OH expression in the daughter cell prevents that cell from undergoing mating-type switching
EXAMPLE 2 Localized mRNAs can establish differential gene expression among the genetically-identical cells of a developing embryo
Just as the fate of the daughter cell is constrained by its inheritance of the ash1 mRNA in yeast, the cells in a developing embryo can be instructed to follow specific pathways of developmentthrough the inheritance of localized mRNA.Such as the cionadevelopment.
This is the Ciona life cycle. They are hermaphrodites and possess both sperm and eggs, then fertilize to develop into a new individual.
Early cleavage in AscidiansThe fertilized, 1-cell ascidian embryo contains a number of localized “determinants” that control the development of different tissues. For example, the yellow determinants is inherited by cells that form the tail muscles.
In the case of muscle differentiation in sea squirts, a major determinant for programming cells to form muscle is a regulatory protein called Macho-1.the Macho-1 mRNA becomes localized in the fertilized egg.
EXAMPLE 3 Cell to cell contact elicits differential gene expression in the sporulating bacterium , B.subtilis.
Under adverse condition,the bacterium Bacillus subtilis can formspores : First, a septum is formed within the sporangium; then the septum produces two cells : forespore (smaller one) and the mother cell (larger one).The mother cell can help the forespore to form a spore. And the forespore can also influences the expression of genes in the neighboring mother cell.
The forespore contains an active form of a specific σfactor,σF (inactive in mother cell). The spollR gene is activated by σF in forespore. The SpollR protein triggers the activation of σE, and leads to the recruitment of RNA polymerase and the activation of specific genes in mother cell.
EXAMPLE 4 A skin-nerve regulatory switch is controlled by notch signaling in the inset CNS
We must first describe the development of the ventral nerve cord in insect embryos. It arises from a sheet of cells called neurogenic ectoderm, then forms two major cell type : neurons and skin cells (or epidermis).
The developing neurons contain a signaling molecule Delta on their surface, which binds to a receptor Notch on the skin cells. The activation of the Notch receptor by Delta renders them incapable of developing into neurons.
The developing neuron does not express neuronal repressor genes. These genes are kept off by a DNA-binding protein called Su(H) and associated repressor proteins. The Delta-notch interactions cause neuronal repressor genes to express.
In all vertebrate embryos, there is a stage when cells located along the future back to move in a coordinated fashion toward internal regions and form the neural tube.Cells located in the ventralmost region of the neural tube form a specialized structure called the floorplate.
Section 3 The molecular biology of drosophila embryogenesis
Development of germ cell Polar granules located in the posterior cytoplasm of the unfertilized egg contain germ cell determinants, and the Nanos mRNA. Nuclei begin to migrate to the periphery. Those that enter posterior regions sequester the polar granules and form the pole cells, which form the germ cells.
Ligation experiment is the first evidence that has shown that cell fate specification is controlled by localized maternal determinants that are deposited into the egg during oogenesis. When a hair is used to separate the anterior and posterior halves of early embryos, then determinants emanating from the anterior pole fail to enter posterior regions. As a result, the embryos develop into abnormal files that lack thoracic structure.
In contrast, when the hair separates older embryos, then the determinant already entered posterior regions and a normal thorax forms.
Embryogenesis of the fruit fly,drosophila melanogaster,has been studied extensively as a relatively simple model for cell differentiation in animals .
During fertilization ,a single sperm cell enters a mature egg ,and the haploid sperm and egg nuclei fuse to form a diploid nucleus.
Overview of Drosophila Development 1.After fertilization, the diploid ,zygotic nucleus undergoes 10 rapid cleavages,the embryo is called a syncitium at this point, because it is a single cell with multiple nuclei. Acturally after 8 cleavages ,the resulting 256 nuclei begin to migrate to the outer edge of the cell. During this migration,the nuclei undergo two more cleavages,resulting 1024 nuclei.
2.About 90 minutes after fertilization, most nuclei have reached the cortex. These nuclei undergo 3 more cleavages
3.From 2-3 hours after fertizilation, cell membrances form between nuclei ,and embryo transforms into a cellular blastoderm.
When the nuclei reach the edge of the cell ,they are totipotent. Just after cellularization,the nuclei have been irreversibly determined to differentiate into specific tissues in the adult fruit fly. The location each nucleus determines its fate.
A morphogen gradient contrals Dorsal-Ventral patterning of the Drosophila embryo This is determinated by a regulatory protein called Dorsal.After fertilization, Dorsal enters nuclei in ventral and lateral regions but remains in the cytoplasm in dorsal regions. Regulated nuclear transport of the Dorsal protein is controlled by a cell signaling molecule called Spätzle. This signal is distributed in a vintral-to-dorsal gradient within the extracellular matrix.
After fertilization, Spätzle binds to the cell surface toll receptor . Toll is activated to a greater or lesser extent by the degree of receptor occupancy in a given region of the syncitial embryo.
Toll signaling causes the degradation of a cytoplasmic inhibitor, Cactus, and the release of Dorsal from the cytoplasm into nuclei.This leads to the formation of a corresponding Dorsal nuclear gradient in the ventral half of the early embryo.
We now consider the regulation of three different target genes that are actived by high,intermediate,and low levels of the Dorsal protein-twist,rhomboid,and sog.
The highest levels of Dorsal protein activate the expression of the twist gene . The reason for this is that the twist 5’regulatory DNA contains two low-affinity Dorsal binding sites.