Download

1 / 16
160 likes | 285 Views
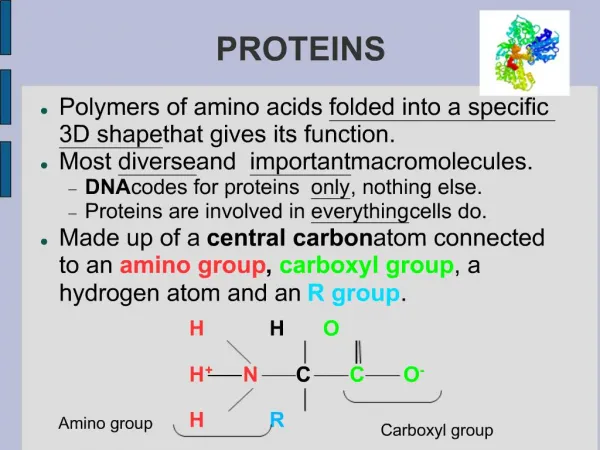
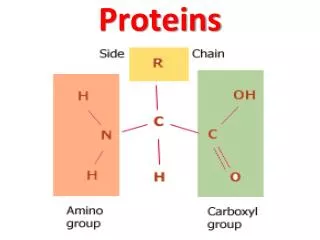
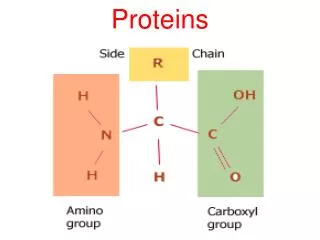
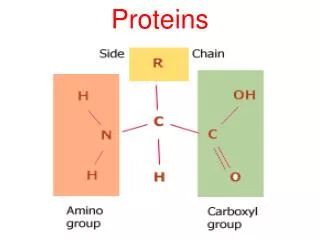
Modular proteins II. Level 3 Molecular Evolution and Bioinformatics Jim Provan. Patthy Sections 8.1.3 – 8.2. Gly. Lys. Val. Asn. Phase 0. GGC AAG. gtaagt ................ (Py) n ncag. GTC AAC. Gln. G. ly. Gln. Phase 1. GG CAA G. gtaagt ................ (Py) n ncag. GT CAA C.

E N D
Modular proteins II Level 3 Molecular Evolution and Bioinformatics Jim Provan Patthy Sections 8.1.3 – 8.2
Gly Lys Val Asn Phase 0 GGC AAG gtaagt ................ (Py)nncag GTC AAC Gln G ly Gln Phase 1 GG CAA G gtaagt ................ (Py)nncag GT CAA C Ala Ar g Ser Phase 2 G GCA AG gtaagt ................ (Py)nncag G TCA AC Intron phase
0 2 1 1 1 2 1 2 1 1 1 0 2 1 1 2 1 2 1 1 1 1 1 1 1 0 2 1 1 2 1 2 1 Intron phase and evolution of collagenases
Exon shuffling by intronic recombination • Middle repetitive sequences flanking an exon may facilitate “looping out” or insertion of modules by intronic recombination • Best example of contraction and expansion of a multidomain protein found in apolipoprotein(a): • Number of tandem kringle domains ranges from 12 to 51 copies • In one variant, 24 of the 37 kringle domains have identical nucleotide sequences, suggesting very recent duplication • Isoforms containing different numbers of kringle domains do not follow simple Mendelian patterns of inheritance: offspring often have apolipoprotein(a) isoforms that differ from those of parents • Such proteins retain interdomain introns: these are responsible for high levels of gene structure plasticity
Factors favouring intronic recombination • Only a tiny portion of spliceosomal introns is essential for splicing: • 5’ end, 3’ end and branch site • Separated by very long sequences that are tolerant to insertions and deletions • Illustrated by comparison of urokinase genes: • Genome organisation of murine, human, porcine and chicken genes is identical in terms of location and phase class of introns • Chicken urokinase introns show hardly any sequence similarity with corresponding mammalian introns, except near splicing junctions and branch sites • Great difference in size of orthologous introns: • Intron A is 1489 bp in chicken • Only 306 bp in humans
Factors favouring intronic recombination • Advantages of spliceosomal introns for exon shuffling (large size, presence of middle repetitive sequences, tolerance to structural changes) holds primarily for vertebrate genomes: • Fungi and plants have fewer and shorter introns • Genes of best studied invertebrate genomes (C. elegans, D. melanogaster) also have shorter introns: • Relatively compact genome may be characteristic of ancestral metazoa • Alternatively, selection may have led to a secondary increase in genome compactness in these lineages • Since plant spliceosomal introns are shorter than those of vertebrates, they are less suitable for intronic recombination • Splicing of chimeric introns, an inevitable consequence of intronic recombination, is impaired in yeast and plants
Acceptance of mutants created by intronic recombination • Several levels of selection determine whether intronic recombination mutant will be fixed or rejected: • Chimeric intron must be spliced correctly, otherwise translation will probably run into a stop codon in the mRNA/intron region and form a truncated protein • Two non-orthologous introns must be in the same phase class: • Must split the reading frame in the same phase • Downstream exon must be translated in its original phase to prevent frameshift mutations • Symmetrical exons • New protein must be able to adopt a stable conformation • Selective advantage of having a new functional domain • Impact of exon insertion may initially be mitigated by alternate splicing
Duplication 2 2 2 2 0 2 2 0 1 2 1 2 0 2 Insertion Deletion 2 1 0 2 Duplication Insertion Deletion The intron-phase compatibilty rule
The symmetrical exon rule • Insertion, deletion and duplication of a module by intronic recombination can satisfy the phase compatibility requirement only if the two introns flanking the module are of the same phase (symmetrical modules): • Only symmetrical module groups are 0-0, 1-1 and 2-2 • Can only be inserted into the compatible intron i.e. 1-1 modules can only be inserted into phase 1 introns • If the structure of a gene of a modular protein conforms to these rules, it suggests that the protein has evolved through exon shuffling
1 1 1 1 1 1 1 1 1 1 2 Factor XIIIb subunit 1 1 1 2 1 1 1 2 1 1 1 2 1 1 1 1 0 2 C3d/Epstein-Barr virus receptor Intron insertion and removal
Evolution of mobile modules • Conversion of a domain to a module: • Protein domain may be converted to a protomodule if introns of identical phase are inserted at its boundaries • Tandem duplication may lead to homopolymerisation • New mobile module may be excised and reinserted at a new location • There is a large variety of class 1-1 modules known, but relatively few class 2-2 or class 0-0 modules • May be due to initial predominance of class 1-1 modules by chance
Example of the modularisation process • The Kunitz type proteinase inhibitor is a single module protein: • In bovine pancreatic trypsin inhibitor gene, phase 1 intons are found at both boundaries of the single inhibitor domain (protomodule stage) • Lipoprotein-associated coagulation inhibitor consists of three tandem copies of this module (tandem duplication stage) – each module is encoded by a distinct class 1-1 exon • Kunitz-type inhibitor modules have been inseted into the genes of other proteins (shuffling stage): • Amyloid precursos • Collagens
Evolution of exon shuffling • Obvious examples of proteins assembled by exon shuffling are restricted to animals: • Not surprising, considering that the evolution of introns and modules is a relatively late development • Recent evolution of spliceosomal pre-mRNA • Large-scale genome projects on model organisms provides information on modular evolution: • Many examples in metazoa – presence of “vertebrate” modules in invertebrates suggests mechanism predates split • No evidence in yeast • Only one possible example in Arabidopsis (receptor protein kinase with two EGF-like domains)
Evolutionary significance of exon shuffling • Number of proteins constructed from modules underlines value of exon shuffling • Several unique features made this mechanism important: • Large collection of binding specificities can coexist in a single protein e.g. plasma proteinases • Acquisition of a new domain can bring about a sudden change in specificity: • Good example is gelatinase • Insertion of a gelatin-binding FN2 module into an ancestral metalloproteinase of the collagen family • Could be correlated with metazoan “big bang”
Modular assembly by exonic recombination • Exon shuffling by intronic recombination may not be the only way to exchange domains between genes: • Modular protein of the bacteria Peptostreptococcus magnus is the product of a recent intergenic recombination of two different types of streptococcal surface proteins • Transfer of one part of a prokaryotic gene to another without the aid of introns • Multidomain bacterial proteins of the PEP sugar transferase system: • Three functional domains separated by “unusual” flexible linker regions • Linkers responsible for frequent rearrangements