Download
1 / 1
10 likes | 147 Views
Interactions of Chemosynthetic Bacteria with Mercury at Deep-sea Hydrothermal Vents Melitza Crespo-Medina 1 , Nicolas Bloom 2 , Aspassia Chatziefthimiou 1 , John Reinfelder 1 , Costantino Vetriani 1 and Tamar Barkay 1

E N D
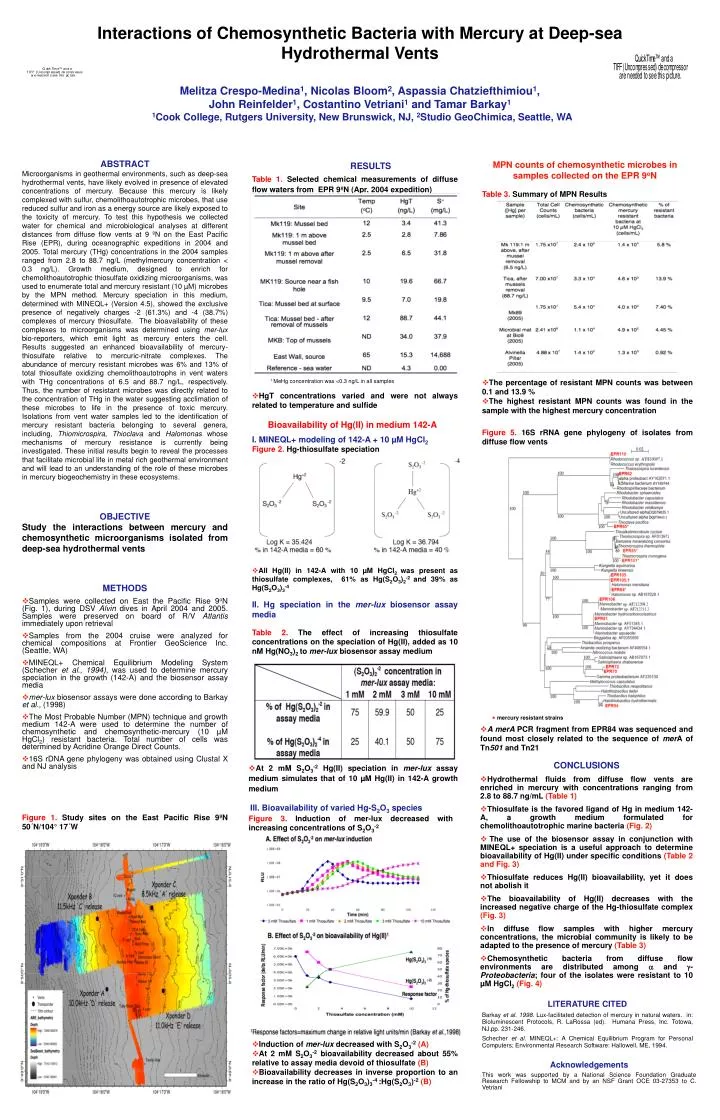
Interactions of Chemosynthetic Bacteria with Mercury at Deep-sea Hydrothermal Vents Melitza Crespo-Medina1, Nicolas Bloom2, Aspassia Chatziefthimiou1, John Reinfelder1, Costantino Vetriani1 and Tamar Barkay1 1Cook College, Rutgers University, New Brunswick, NJ, 2Studio GeoChimica, Seattle, WA ABSTRACT Microorganisms in geothermal environments, such as deep-sea hydrothermal vents, have likely evolved in presence of elevated concentrations of mercury. Because this mercury is likely complexed with sulfur, chemolithoautotrophic microbes, that use reduced sulfur and iron as a energy source are likely exposed to the toxicity of mercury. To test this hypothesis we collected water for chemical and microbiological analyses at different distances from diffuse flow vents at 9 ºN on the East Pacific Rise (EPR), during oceanographic expeditions in 2004 and 2005. Total mercury (THg) concentrations in the 2004 samples ranged from 2.8 to 88.7 ng/L (methylmercury concentration < 0.3 ng/L). Growth medium, designed to enrich for chemolithoautotrophic thiosulfate oxidizing microorganisms, was used to enumerate total and mercury resistant (10 µM) microbes by the MPN method. Mercury speciation in this medium, determined with MINEQL+ (Version 4.5), showed the exclusive presence of negatively charges -2 (61.3%) and -4 (38.7%) complexes of mercury thiosulfate. The bioavailability of these complexes to microorganisms was determined using mer-lux bio-reporters, which emit light as mercury enters the cell. Results suggested an enhanced bioavailability of mercury-thiosulfate relative to mercuric-nitrate complexes. The abundance of mercury resistant microbes was 6% and 13% of total thiosulfate oxidizing chemolithoautotrophs in vent waters with THg concentrations of 6.5 and 88.7 ng/L, respectively. Thus, the number of resistant microbes was directly related to the concentration of THg in the water suggesting acclimation of these microbes to life in the presence of toxic mercury. Isolations from vent water samples led to the identification of mercury resistant bacteria belonging to several genera, including, Thiomicrospira, Thioclava and Halomonas whose mechanisms of mercury resistance is currently being investigated. These initial results begin to reveal the processes that facilitate microbial life in metal rich geothermal environment and will lead to an understanding of the role of these microbes in mercury biogeochemistry in these ecosystems. MPN counts of chemosynthetic microbes in samples collected on the EPR 9oN RESULTS Table 1. Selected chemical measurements of diffuse flow waters from EPR 9ºN (Apr. 2004 expedition) Table 3. Summary of MPN Results 1 MeHg concentration was <0.3 ng/L in all samples • The percentage of resistant MPN counts was between 0.1 and 13.9 % • The highest resistant MPN counts was found in the sample with the highest mercury concentration • HgT concentrations varied and were not always related to temperature and sulfide Bioavailability of Hg(II) in medium 142-A Figure 5. 16S rRNA gene phylogeny of isolates from diffuse flow vents I. MINEQL+ modeling of 142-A + 10 µM HgCl2 Figure 2. Hg-thiosulfate speciation OBJECTIVE Study the interactions between mercury and chemosynthetic microorganisms isolated from deep-sea hydrothermal vents • All Hg(II) in 142-A with 10 µM HgCl2 was present as thiosulfate complexes, 61% as Hg(S2O3)2-2 and 39% as Hg(S2O3)3-4 • METHODS • Samples were collected on East the Pacific Rise 9oN (Fig. 1), during DSV Alvin dives in April 2004 and 2005. Samples were preserved on board of R/V Atlantis immediately upon retrieval • Samples from the 2004 cruise were analyzed for chemical compositions at Frontier GeoScience Inc. (Seattle, WA) • MINEQL+ Chemical Equilibrium Modeling System (Schecher et al., 1994), was used to determine mercury speciation in the growth (142-A) and the biosensor assay media • mer-lux biosensor assays were done according to Barkay et al., (1998) • The Most Probable Number (MPN) technique and growth medium 142-A were used to determine the number of chemosynthetic and chemosynthetic-mercury (10 µM HgCl2) resistant bacteria. Total number of cells was determined by Acridine Orange Direct Counts. • 16S rDNA gene phylogeny was obtained using Clustal X and NJ analysis II. Hg speciation in the mer-lux biosensor assay media Table 2. The effect of increasing thiosulfate concentrations on the speciation of Hg(II), added as 10 nM Hg(NO3)2 to mer-lux biosensor assay medium * mercury resistant strains • A merA PCR fragment from EPR84 was sequenced and found most closely related to the sequence of merA of Tn501 and Tn21 CONCLUSIONS • Hydrothermal fluids from diffuse flow vents are enriched in mercury with concentrations ranging from 2.8 to 88.7 ng/mL (Table 1) • Thiosulfate is the favored ligand of Hg in medium 142-A, a growth medium formulated for chemolithoautotrophic marine bacteria (Fig. 2) • The use of the biosensor assay in conjunction with MINEQL+ speciation is a useful approach to determine bioavailability of Hg(II) under specific conditions (Table 2 and Fig. 3) • Thiosulfate reduces Hg(II) bioavailability, yet it does not abolish it • The bioavailability of Hg(II) decreases with the increased negative charge of the Hg-thiosulfate complex (Fig. 3) • In diffuse flow samples with higher mercury concentrations, the microbial community is likely to be adapted to the presence of mercury (Table 3) • Chemosynthetic bacteria from diffuse flow environments are distributed among and - Proteobacteria;four of the isolates were resistant to 10 µM HgCl2(Fig. 4) • At 2 mM S2O3-2 Hg(II) speciation in mer-lux assay medium simulates that of 10 µM Hg(II) in 142-A growth medium III. Bioavailability of varied Hg-S2O3 species Figure 1. Study sites on the East Pacific Rise 9ºN 50´N/104° 17´W Figure 3. Induction of mer-lux decreased with increasing concentrations of S2O3-2 LITERATURE CITED Barkay et al. 1998. Lux-facilitated detection of mercury in natural waters. in: Bioluminescent Protocols, R. LaRossa (ed). Humana Press, Inc. Totowa, NJ.pp. 231-246. Schecher et al. MINEQL+: A Chemical Equilibrium Program for Personal Computers; Environmental Research Software: Hallowell, ME, 1994. • Induction of mer-lux decreased with S2O3-2(A) • At 2 mM S2O3-2 bioavailability decreased about 55% relative to assay media devoid of thiosulfate (B) • Bioavailability decreases in inverse proportion to an increase in the ratio of Hg(S2O3)3-4 :Hg(S2O3)-2(B) Acknowledgements This work was supported by a National Science Foundation Graduate Research Fellowship to MCM and by an NSF Grant OCE 03-27353 to C. Vetriani