Download

1 / 62
660 likes | 1.04k Views
MHC . A Review. MHC. Originally discovered in the 1930s by Peter Gorer ( gorer , 1936) MHC was genetically defined by George Snell (1948) Human Leukocyte Antigen (HLA) was sequences by Betck et al. in 1999

E N D
MHC A Review
MHC • Originally discovered in the 1930s by Peter Gorer (gorer, 1936) • MHC was genetically defined by George Snell (1948) • Human Leukocyte Antigen (HLA) was sequences by Betck et al. in 1999 • MHC is a complex multigene family comprising loci encoding receptors on the surface of a variety of immune and nonimmune cells • These receptors bind amino acid fragments (or peptides) from foreign pathogens to start an immunological response
MHC • The diversity of MHC genes underlies resistance to the diverisyt of infectious pathogens (Zuckerkandle and Pauling) • “Classical” is used to indicate that an MHC gene is polymorphic and highly expressed • “Nonclassical” genes are MHC genes that cluster with other MHC genes phylogenetically • They are usually not expressed at high levels and are not polymorphic
General Organization and Inheritance of MHC • Every mammalian species studied possesses a tightly linked cluster of genes, the major histocompatibility complex (MHC) • Associated with • Intercellular recognition • Self/nonself discrimination • MHC plays a central role in the development of both humoral and cell-mediated immune responses
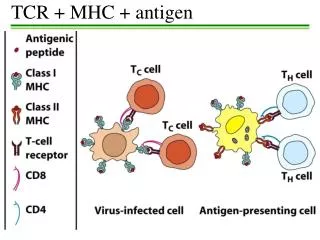
General Organization and Inheritance of MHC • Role in antigen recognition by T-cells • Antigen-presenting structures • TH and Tc cells • Partly determines the response of an individual to antigens of infectious organisms and the MHC has been implicated in the susceptibility to disease and in the development in autoimmunity
Location and Function of MHC Regions • Chromosome 6 in humans = HLA • The most gene-rich region, with over 200 loci identified • Chromosome 17 in mice = MHC/ H-2 • 3 classes of molecules • Class I MHC genes • Encodes glycoproteins expressed on the surface of nearly all nucleated cells • Present peptide antigens of altered self-cells necessary for the activation of Tc cells
Class I MHC genes • Encodes glycoproteins expressed on the surface of nearly all nucleated cells • Present peptide antigens of altered self-cells necessary for the activation of TC cells • Encoded by the A, B and C regions in humans
Class II MHC genes • Encodes glycoprteinsexpressed primarily on antigen-presenting cells • Macrophages, dendritic cells and B cells • Presents antigenic peptides to TH cells • Encoded by DP, DQ and DR regions in humans
Class III MHC genes • Encodes secreted proteins associated with the immune process including soluble serum proteins, components of the complement system and tumor necrosis factor • C4, C2, Bf (complement proteins) in humans
MHC • Individuals inherit the alleles as two sets, one from each parent. • Each set of alleles if referred to as a haplotype • In an outbred population of humans the offspring are generally heterozygous at many loci and will express both maternal and paternal MHC alleles • Co-dominantly expressed on the same cell
Generations • If two inbred strains of mice having different MHC haplotypes are bred, the F1 generation inherits haplotypes from both parental strains and expresses both parental alleles at each MHC locus • Congenic Mice = two strains that are genetically identical except at a single locus or region • The first letter in a congenic strain refers to the strain providing the genetic background and the second letter to the strain providing the genetically different MHC region.
MHC Molecules and Genes • Class I and Class II MHC molecules are membrane-bound glycoproteins • Antigen presenting molecules • Class II MHC molecules are soluble proteins and exhibit much less polymorphism than class I and II
Structure of Class I Molecule • Contains a large α chain associated noncovalently with the much smaller β2-microglobulin molecule. • The α chain is a polymorphic transmembraneglycoproteins • 45 kilodaltons (kDa) • Encoded by genes within the A, B and C region of the HLA in humans • Anchored in the membrane by hydrophobic transmembrane segment and hydrophilic cytoplasmic tail • β2-microglobulin is an invariant protein of about 12kDa encoded by a gene located on a different chromosome
Structure of Class I Molecule • α chain molecules are organized into three external domains (α1, α2, α3) each containing approximately 90 amino acids • A transdomain of about 40 amino acids and a cytoplasmic anchor segment of 30 amino acids • Β2-microglobulin is similar to α3 external domain • Papain (enzyme) cleaves the α chain 13 residues proximal to its transmembrane domain
Structure of Class I Molecule • α1 and α2 domains interact for form a platform of eight antiparallelβ strands spanned by two long α-helical regions • This structure forms a deep groove or cleft approximately 25 Å X 10 Å X 11Å • The long α helices as sides and the β strands of the β sheet as the bottom • The Peptide Binding Cleft is located on the top surface of the class I MHC molecule • Can bind 8-10 amino acids
Peptide Binding by MHC Molecules • Several hundred different allelic variants of class I MHC molecules in humans • Each individual expresses only up to 6 different class I molecules and up to 12 different class II molecules • A given MHC molecule can bind numerous different peptides • Some peptides can bind to several different MHC molecules • Class I molecules bind peptides containing 8 – 10 amino acid resides
Class I MHC-Peptide Interaction • Class I MHC molecules bind peptides • Present these to CD8+T cells • A nucleated cell expresses about 105 copies of each class I molecule • Many different peptides will be expressed simultaneously on the surface of a nucleated cell by class I MHC molecules • Class I MHC can bind over 2,000 distinct peptides • Presented with a frequency of 100-4,000 copies per cell
Polymorphism of Class I Molecules • Polymorphism – Multiple alleles at a given genetic locus within a species • MHC expressed by an individual does not change over time • One of the most polymorphic genetic complexes • Possesses an extraordinarily large number of different alleles at each locus • The alleles differ in their DNA sequences from one individual to another • The gene products expressed by different individuals have unique structural differences
Polymorphism of Class I Molecules • Up to 20 amino acid residues contributing to the uniqueness of each allele • Analysis of HLA class I molecules has so far revealed 59 A alleles, 111 B alleles, and 37 C alleles. • The current estimate of actual polymorphism in humans is on the order of 100 alleles for each locus • The binding site for antigens is the most polymorphic location on MHC Class I
MHC Genes • Chromosome 6 in humans and 17 in mice • Nearly 100 genes spanning some 4000kb • 2000kb for Class I • Contains approximately 20 genes • HLA-A, HLA-B, HLA-C • HLA-A has 250 and HLA-B has 500 known alleles • Non-classical MHC class I include: HLA-E, HLA-F, HLA-G, HLA-H, HLA-J, HLA-X and MIC • Many of these are pseudogenes and do not encode a protein product • The non-classical gene products are categorized as class Ib molecules
Class Ib Molecules • Have an α chain that associates with β2-microglobulin • Less polymorphic • Expressed at lower levels • Tissue distribution is more limited than classical class I molecules • The function of non-classical class I MHC molecules is largely unknown
Classical Class I MHC Molecules • Expressed on most somatic cells • Level of expression varies • Highest level of expression are by lymphocytes • 1% of the total plasma-membrane proteins, 5X105 molecules per cell
Regulation of MHC Expression • Interferons (alpha, beta and gamma) and tumor necrosis factor increase expression of class I MHC molecules on cells • MHC expression is increased or more commonly, decreased by a number of viruses, ie. Cytomegalovirus, hepatitis B virus and adenovirus 12.
Chemosensationand Genetic Individuality • Olfactory cues play a fundamental role is social interactions in a wide variety of mammals from rodents to primates (Brown, 1979) • Each mammal has an individual odour or signature for individual recognition (Porter, 1985) • Olfactory signatures have a function in mammalian reproduction (MacDonald and Brown, 1985) • Chemical signals can be deposited on external substrates to mark territory • Olfactory cues can still be effective in darkness (Porter, 1999)
Immunobiology of Classical MHC class I antigens • Outbreeding mammals have a degree of polymorphism that ensures that in each species there are more than 109 unique class I antigenic phenotypes in a population • The heavy chain consists of three domains (α1, α2 and α3) • The light chain consists of a single domain, β2-microglobulin, attached non-covalently to the α3 domain
Immunobiology of Classical MHC class I antigens • The α1 and α2 domains consist of an α-helix and a β-pleated sheet. • The two sheets join together to make a continuous platform which supports the two α-helices defining an antigen binding cleft • The cleft is occupied by a self-peptide of 8-10 amino acids
MHC-associated Odours and Individual Recognition • Inbred strains consist of animals that are genetically uniform and homozygous for all their genes • Each chromosome pair is identical • Resulting from mating brother and sister pairs for at least 20 consecutive generations • The genotype is nearly identical • Congenic strains are two strains that are identical except for allelic differences at a particular locus • Any measurable characteristic that differs between the two congenic strains must be ascribed to the genes at that locus • Formed by repeated backcrosses of a donor strain to an inbred strain with selection for a locus of interest
The Y-maze Paradigm • Mice are trained to enter alternative chambers scented by an airflow through odour boxes occupied by MHC-congenic mice • Mice are trained by water deprivation and reward • The mouse first learns to differentiated between two strong odours like juniper and cinnamon and is rewarded with a drink • It is correct if it chooses the correct arm at least 80% of the time • The mice learn to distinguish inbred strains and then distinguish congenic strains that differ only in the alleles at the MHC
The Y-maze Paradigm • Findings: • No sensory perception other than olfaction is required to distinguish between MHC types (Yamazaki, 1979) • Urine is a good source of MHC-specific odours (Yamaguchi, 1981) • Males and females had equal ability to distinguish between MHC types (Yamaguchi, 1981) • Olfactory acuity of mice could enable the discrimination of MHC Class I molecules that differ only in three amino acids (Yamazaki, 1983a) • No amount of training could enable a mouse to distinguish between individuals of the same inbred strain (Yamaguchi, 1981)
The Y-maze Paradigm • Mice could discriminate between allelic differences in the class II and Q/T/M regions, indicating that all three genetic loci of the MHC can contribute to individual odours (Yamazaki, 1982, 1984) • The haemopoietic system is a source of MHC-associated odours (Yamazaki, 1985) • X and Y chromosomes are also discriminated between, but are much less potent in scent marking than the MHC (Yamazaki, 1986a) • Ease of training indicates that MHC is a potent source of individual odour (Boyse, 1987)
Habituation-Dishabituation Paradigm • Most mammal are usually inquisitive when presented with a novel stimulus, but the novelty can wear off upon extended exposure to the stimulus • The test involves placing an experimental animal in a test arena with a removable wire lid • The animal is given opportunity to explore • Then it is presented with the first urine odour on a piece of filter paper attached to the wire lid • A novel stimulus
Habituation-Dishabituation Paradigm • The animal rears on its hind legs and sniffs • The urine sample is presented three times • By the 3rd exposure it no longer rears to investigate • It has habituated to the odour • A novel urine sample is presented to the animal • If it perceives the odour of the new sample to be different from the first there is a rapid dishabituation • The rat lifts its head, sniffs the air and moves toward the new odour and rears to investigate it
MHC-associated odours and reproduction • The male will choose a female that is different from him at the MHC (Yamazaki, 1976) • Females can also choose mates and prefer to mate with males of different MHC (Potts, 1991) • “Bruce Effect” – the pregnant female is introduced to a strange male during the preimplantation stage of development and spontaneous abortion occurs (Yamazaki, 1983b) • The effect is likely due to a neuroendocrine disturbance
MHC-associated odours: the “carrier hypothesis” • MHC-based developmental variations give rise to distinctive odour profiles (Boyse, 1987) • It is possible to discriminate between urine samples taken from germ-free MHC congenic mice, which lack commensal flora (Yamazaki, 1990) • Fetal MHC-associated odours are evident in the urine of mothers as early as day 9 of gestation (Beauchamp, 1994) • Before the fetus has a functioning immune system (Owen, 1969)
MHC-associated odours: the “carrier hypothesis” • A hypothetical route of class I MHC molecules from the membrane to the circulation and into the urine • Cleavage of the molecule at the juxta-membranous protease site removes the transmembrane region and releases the molecule into the circulation in soluble form • The molecule does not aggregate and circulates as a soluble heterodimer consisting of α and β chains • Further enzymatic cleavage at the junction of the α2 and α3 domains removes most of the 3 domain with the attached β2-microglobulin • This cleavage allows relaxation of the binding platform, opening of the cleft and loss of bound peptide
MHC-associated odours: the “carrier hypothesis” • Further enzymatic cleavage at the junction of the α2 and α3 domains removes most of the 3 domain with the attached β2-microglobulin • This cleavage allows relaxation of the binding platform, opening of the cleft and loss of bound peptide • The resulting molecule with a naked binding cleft is capable of binding a cocktail of small aromatic molecules such as volatile-odorants and is small enough to pass into the urine
MHC-associated odours: the “carrier hypothesis” • In the serum the molecule is a heterodimer with a heavy chain of 39kDa associated non-covalently with β2-microglobulin • Present in the serum as a concentration between 350 – 390 ng/ml • A half-life of 2.7 h • Excreted via the kidneys • In the urine degradation occurs giving rise to a major molecular mass species of 27kDa • Cleavage at the papain cleavage site
MHC-associated odours: the “carrier hypothesis” • Class I molecules are large and consequently lack a vapour pressure • Class I molecules might associate with smaller molecules, in an allelic-specific way, and transport them from the blood into the urine • Serum has an odour but cannot be used to discriminate between MHC types (Brown, 1987, Yamazaki, 1999) • Treat with proteases to liberate odour • Carboxylic acids may be a source of the odour
In Search of the Chemical Basis for MHC Odourtypes (Kwak, 2010) • Lewis Thomas (1975) • Two different classes of compounds have been proposed: • Small volatile molecules and non-voltaile MHC peptide ligans (Boehm, 2005, Restrepo, 2006) • Volatile molecules are • non-ionic and • have a finite vapour pressure between ambient and 250°C • Their molecular weights are generally less than 300Da
Odourtype • Volatile molecules are • non-ionic and • have a finite vapour pressure between ambient and 250°C • Their molecular weights are generally less than 300Da • MHC peptide ligands are • Ionic and • Composed of inne amino acids • Their molecular weights exceed 1000Da
Chemical Investigations of MHC Odourtype in Mice • Mice can recognize the MHC-driven odour cues from urine and blood • Studies have failed to find specific behavioral effects of MHC variation in wild mice (Cheetham, 2007, Sherborne 2007, Thom 2008, Potts 1991)
Chemical Investigations of MHC Odourtype in Mice • Using anion exchange chromatography confirmed that acidic compounds were responsible for the MHC odour cues • Some voltaile organic acids are regulated by MHC genetic variation (Singer, 1997) • GC/MS study of about 80 compounds associated with MHC (Willse, 2005) • MHC associated odour cue can be recognized in spite of variation in background strains (Beauchamp, 1990, Yamazaki, 1994, Eggert, 1996, Willse, 2006)
Chemical Investigations of MHC Odourtype in Mice • MHC-regulated metabolic odour signal must have a background-independent component • GC/MS experiments on 3 MHC-congenic mice derived from different background strains • 36 out of 55 peaks from urine were found to be present in all three strains • Background genes, MHC genes and their interaction regulate the urinary volatile profiles • SPME GC/MS on mice (Kwak 2009)
Chemical Investigations of MHC Odourtype in Mice • A compound was found to be more prominent in one MHC type in one strain and less prominent in that same MHC type in the other strain • MHC regulation of volatile metabolic phenotypes was modulated by background genes • All published chemical investigations have failed to find qualitative differences in volatile compounds that are associated with MHC types, except Eggert, 1996 • The patterns of volatile metabolites vary according to MHC types, inconsistently and in a complex way
The Major Histocompatibility Complex and the Chemosensory Signaling of Individuality in Humans • Four characteristics of MHC is responsible for its enormous diversity • A number of functional loci per class • Allelopolymorphism (estimated for HLA class I raging up to 60 different alleles per locus) • Codominant expression • A high level of heterozygocity (in natural populations of mice, it is almost 100%)
The Major Histocompatibility Complex and the Chemosensory Signaling of Individuality in Humans • MHC effects on social behavior in humans came from three types of studies • A series of population studies on the occurrence of shared HLA types in couples which in part confirms an influence of the MHC but are not conclusive • Experimental studies on trained rats’ responses to human urinary odor which demonstrated preliminary results of an MHC-associated odor expression in humans • A field study in which women were asked to index others whose body odors elicited a strong response • A higher communality between indexing females and indexed females than with indexed males • Males for whom negatively rated body odors were reported show a higher genetic similarity than those for whom positively rated body odors were reported (Eggert, 1994)
Investigate trained rats’ responses to human odors • Rats are able to distinguish between the urine odors of immunogenetically defined HLA-homogenous groups of persons • HLA-associated urine donors occur in members of both sexes • Stimulus generalization task can be performed using urine donors with different HLA-types • HLA-associated odor constitution appears not to be limited to a definite HLA-specificity
Fractionation and Bioassay of Human Odor Types • Which fraction in human urine are chemosensorically active? • The role of fractions containing either volatile or nonvolatile substances • Bioassay using rats sniffing volatiles, smaller than 250Da and nonvolatiles • Rats were able to recognize the odor profile of the training samples in the fractionated residue • The distillate from the urine samples containing the volatile urine fraction was not recognized
GC of Human Urine • Using a GC on human urine 120 peaks were found • 16 out of 120 of specific components associated with the examined HLA-types were found • HLA differences influence the pattern of ubiquitous metabolites • Is the body odor of humans associated with HLA-types? • The constitution of body odors in humans may provide evidence of an ultimate function of MHC-associated odors in humans
Fractionation and Bioassay of Human Odor Types • All tests showed that the bioactive information of the odor profile is not included in the distillate containing only the volatiles. • Olfactory information was not changed by adding a foreign distillate to a familiar residue • Olfactory recognition only occurred when the residue of the fractionated urine samples were presented • Odor conveyed by the volatiles disappears very quickly. • The mechanism of slowly released volatiles would prevent a rapid disappearance of the individual scent