Download

1 / 27
270 likes | 273 Views
Biology of Dicosmoecus gilvipes in Northern California Streams. Vincent H. Resh, Morgan Hannaford, John Jackson, and Patina Mendez Department of Environmental Science Policy & Management University of California, Berkeley. Dicosmoecus gilvipes (Hagen).

E N D
Biology of Dicosmoecus gilvipes in Northern California Streams Vincent H. Resh, Morgan Hannaford, John Jackson, and Patina Mendez Department of Environmental Science Policy & Management University of California, Berkeley
Dicosmoecus gilvipes (Hagen) • Occurs from Northern California to British Columbia and east to Montana, Nevada, Idaho and Wyoming • Large size, high densities,synchronous emergence make it important in trout diets • To anglers it’s the “Giant Orange sedge”, “October Caddis”
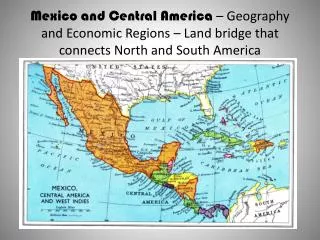
Study Sites in Northern California • McCloud River—larval behavior • Eel River—phenology and grazing studies • Big Sulphur Creek—adult behavior
Egg Masses • Found at the base of Carex sedges • Not found on leaves overhanging streams as reported for some other Dicosmoecinae (after hundreds of hours of searching) ?
Larval Cases • Early-instar cases made from organic material and detritus • With 3rd instar, some pebbles incorporated, a time when discharge begins to recede (except in snow-melt streams) • 4th instar increases stone material in case; 5th instar cases entirely made of stones (30mm in length)
Larval Biology • Larvae are highly mobile moving up to 25 m/day From Hart and Resh 1980
Among first benthic macroinvertebrates to colonize storm-scoured, mid-channel areas • Predominantly scrapers-grazers: gut content contain diatoms, filamentous algae, and some mineral particles • Densities 80-100 larvae/m2
Developmental Diapause and Pupation • 5th instars attach cases to undersides of boulders and remain dormant until pupation • Diapause lasts one month, followed by pupation • Pharate adults emerge from cases on underside of boulders and crawl to shore in late afternoon
Temperature influences on Development Average accumulated degree days (>0 C) to reach fifth instar: • Upstream-most site, 2,485 degree days • 4 km downstream, 2,585 degree days D. gilvipes at downstream site reach 5th instar and diapause 2 weeks earlier than at upstream site • D. gilvipes is rare below the downstream site, perhaps reflecting temperature tolerance
Phenology • In California streams, D. gilvipes exhibits a univoltine life cycle (add graph) • In Flathead River in Montana, 2-yr cycle (Hauer & Stanford 1982) • 1st instars appear in spring and grow through summer, overwintering as 5th instars in apparent diapause • 5th instars are active again when temp >1C, and then enter 4-6 wk prepupal diapause, emerging August to October
Adults • Adult flight and mating occur in early evening • Pheromone-based mating system
Pheromone Studies • Live females of D. gilvipes attract males during the night with 58% being captured within 1 hr after sunset • Timing of male response to female pheromone was the same as male response to live females • Males and females fly during periods of mate attraction but not generally at other times • Males but not female of D. gilvipes exhibit circadian rhythms • The brief but coordinated activities may contribute to both reproductive success and reproductive isolation • Pheromones are produced by paired exocrine glands located on 5th sternite Resh et al. 1987; Jackson and Resh 1991
Behavioral Studies in the Laboratory • Time spent in feeding, crawling, resting, and case building varied among 3rd, 4th, and 5th instars (Li and Gregory 1989) • Larvae feed ½ time, 3rd instars fed longer on diatoms, 4th instars longer on filamentous algae • 4th instars spend most time case building; 5th instars resting • Neither aggression nor cannibalism were observed
Behavioral Studies in the Field • Patterns of movement of D. gilvipes follow a random-walk model except large larvae move farther than smaller larvae, and there is a pronounced diel rhythm to movement (traveling faster during the day) (Hart and Resh 1980) • 2/3 of time budget spent in feeding (Hart 1981) • Larvae feed in areas with more food, and minimize “turn angle”, reducing possibility that they return to a recently grazed patch • In some streams, larvae may compete exploitatively for food • Reflecting their food sources, D. gilvipes can account for 55-96% of biomass in exposed- riparian stream sections (Tait et al. 1994)
Dicosmoecus gilvipes as Grazers: Laboratory Studies • D. gilvipes drastically reduced periphyton biomass and resulted in a shift to a blue green algae community (Walton et al. 1995) and grazed entire substrates cf. the patchy feeding of Juga snails or Baetis mayflies (Dennicola et al. 1990) • D gilvipes doesn’t completely remove algal assemblage but changes growth form and alfgal assemblage physiognamy (Steinman et al. 1987) • D. gilvipes grazing effects was higher than Juga snails or Baetis mayflies (Lamberti et al. 1987, 1992, 1995) • Growth of D. gilvipes is density dependant and highest at its lowest densities (Lamberti et al. 1995) • D. gilvipes has high growth rates but low efficiency in resource use (cf. Juga) • Different life history strategies may explain coexistence of different grazers Juga and D. gilvipes
D. Gilvipes Substrate relationships • Sediment size has significant effects on D. gilvipes survival; 3rd instar more sensitive than 5th instars most (Parker et al. 2002) • D. gilvipes show that sampling efficiency can be optimized because non-aggregated spatial distribution patterns can occur in uniform substrate patches (Lamberti and Resh 1979) • This study underscored the value of habitat stratification
Dicosmoecus gilvipes as Prey • In late June and July, D. gilvipes are 80% of Harlequin ducks (Histrionicus histrionicus) diets (in Oregon, Wright et al. 2000) • After a severe flood, numbers of D. gilvipes were reduced and numbers of harlequin duck numbers were reduced and brood development was delayed • D. gilvipes make up a significant component of American dipper (Cinclus mexicanus) and D. gilvipes inhabit deeper pools in response to dipper predation (in Utah, Hart and Marti 1993; in California, Teague et al 1985) • Human recreational activities effect early instars more than later instars of D. gilvipes (in Oregon, Wright and Li 1998) • D. gilvipes may transmit trematodes that when ingested by horses cause Potomac Horse Fever (Madigan et al. 2000)
Interactions With Other Organisms—Study Design • 18 0.8-m2 cylinder cages used • Exclusion of D. gilvipes • Enclosures (40 D. dilvipes/m2) • Ambient density controls (allowed D. gilvipes access • Ambient densities measured to assess effect of cages on providing D. gilvipes access
Biotic Effects • Fewer sessile invertebrates colonized D. gilvipes enclosures than either in exclosures or controls • A negative correlation existed between densities of active D. gilvipes and densities of sessile organisms • Algal biomass significantly lower in D. gilvipes enclosures than in exclosures or controls • D. gilvipes functions as a “keystone species” in the Eel River
Number of Sessile Organisms at Different Densities of D. gilvipes Tinodes Petrophila Antocha Cricotopus Polycentropus Tanytarsus Simulium
Direct Interference of D. gilvipes with Sessile Grazers--Tinodes • 30x15 cm boxes stocked with cobbles containing Tinodes (Psychomyiid caddisfly) larvae and tube cases • 2 D. gilvipes larvae added to each box • % occupancy of tubes before and after addition used to determine effect of addition • Add slides of Tinodes
Mediterranean Climates Can Influence Results of Field Experiments 18 yrs of field observations and 5 yrs of experiments in the Eel River, California, by Mary Power Inferences about food webs differ during flood and drought years Number of trophic levels connecting algal biomass to consumers differ in a given year Strong biotic interactions differ in a given year Behavioral trophic cascades differ in a given year
Why Do Food Webs Differ Year-to-Year? • Cladophora algal or diatom blooms are basis of food chain • Trout suppress grazing insect populations during years with floods and algae increase • BUT if drought occurs, D. gilvipes are more abundant and Cladophora biomass is lower • Slides of Cladophora, diatoms, steelhead, dicosmoecus, Baetis
Oddest Relationship • D. gilvipes may transmit trematode worms that when ingested by horses cause Potomac Horse Fever (Madigan et al. 2000) • Horses were placed in water with cercariae, drank this water, and were fed insects infected with metacercaria • The parasite is maintained in aquatic ecosystems and can occur through accidental ingestion of Dicosmoecus!