Download

1 / 17
170 likes | 174 Views
Modeling hybridization kinetics. Jian-Ying Wang, Karl Drlica Mathematical Biosciences 183 (2003) 37-47. Introduction (1). DNA-RNA & RNA-RNA hybridization are important : nucleic acid (n.a.) func’n, detection of n.a., alteration its exp’n Two parameters that characterize the hyb.

E N D
Modeling hybridization kinetics Jian-Ying Wang, Karl Drlica Mathematical Biosciences 183 (2003) 37-47
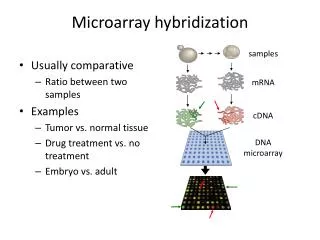
Introduction (1) • DNA-RNA & RNA-RNA hybridization are important : nucleic acid (n.a.) func’n, detection of n.a., alteration its exp’n • Two parameters that characterize the hyb. • Equilibrium yield (main concern when incubation time is unlimited, e.g. Southern hyb.) • Annealing rate (when hyb. is involved in the inhibition of gene exp’n)
Necessary to use experimental methods to determine favorable sites for antisense attack of RNA Introduction (2) • Effective antisense oligont. for hyb. to target RNA • Previous efforts at computational identification • Predictions of RNA 2° structure ( identify regions likely to be single stranded & presumably accessible for hyb. ) -> correlation w/ antisense oligodeoxynt. (ODN) hyb. Shows considerable scatter. ( ss regions don’t correspond to regions that hybridize most readily) • Calculate overall E gain due to hybrid formation & relate to inhibition of gene exp’n -> do not always correlates with experimental data. -> more problematic is the inability of E gain calculations to explain the large (up to 1000 fold ) difference in hyb. rate
Introduction (3) • Important factors in determining accessibility for hyb. : melting E & E (free E) gain • develop a computational description for hyb. of short complementary regions within large n.a. • calculate a rate factor (that is proportional to the rate at which oligont. hybridize to sites in RNA under quasi st-st conditions) rate factor model
Theoretical Considerations • A : antisense oligonucleotide • S : target RNA • AS : antisense-target RNA hybrid • P : products of the second step • The second step may be : an enzymatic cleavage (e.g. antisense-DNA mediated RNase H cleavage rxn ) : structural change ( e.g. annealing of long antisense-target RNA hyb. )
by kinetic approach of quasi st-stconditions, d[AS]/dt=0 thus, the 2nd order association constant : for the initial rate of a single-tube assay, total antisense is conserved, [A0]=[A]+[AS] (at beginning, [P]~0) thus, the 2nd order association constant : • Estimation of Km enables us to compare relative rxn rates • for different sites in a given RNA ( if, k2 ~constant )
1 • to estimate Km, first consider eq. constant for hybrid dissociation • from Boltzmann distribution, the occupancy of the activated state is proportional to • k1* : forward rate constant for the binding of the activated antisense oligont to the activated target site : related to assay conditions, independent of RNA structure • k2/k1* can be considered constant
1 • Can be used for compare different sites in RNA • for antisense ODN hyb. • For this, define a rate factor, x=k/k2 for comparing relative hyb. rate at st-st for estimation of initial hyb. rate (pre-equil.)
Correlations bt’n rate factor & experimental antisense-mRNA hyb. determined w/ oligonucleotide libraries • As a test for • 기존 실험 결과와 비교. • hyb. in the presence of RNase H. • the most readily hyb. regions -> degraded by RNase H • and revealed by gel electrophoresis
Correlation bt’n maximal rate factor and initial hyb. rate • For oligont. longer than 15 nt. • by considering all 15 nt region, & using , calculate maximum rate factor. • The maximal rate factor shows good correlation w/ the 2nd order binding rxn rate constant. ( Accessibility score is based on a statistical consideration of single-strandedness. )
Discussion • An expression was derived that relates nucleic acid secondary structure to the steady-state rate of oligont.–RNA hyb. • The assumption of the model : hyb. of nucleic acids occurs through an intermediate state • The central feature of the rate factor model is that it uses both melting energy and energy gain to identify favorable sites for hyb. • Since dynamic processes are more likely to be relevant than eq. processes for inhibition of gene expression with antisense oligonucleotides, the st-st model should be more suitable than eq. models for identifying favorable hyb. sites.
Materials and methods • G0anti and G0target were obtained from the output files of the GCG RNA folding program FoldRNA (GCG Package Version 10-Unix, Genetics Computer Group, University of Wisconsin, Madison, WI)
extra discussion… • DNA / RNA hyb. rate constant data는 많은데, DNA / DNA는 잘 못찾겠음… • DNA / DNA hyb. rate constant에 대한 experimental data가 있으면, modeling한 equation으로 각 strand fragment의 concentration 추이를 알아볼 수 있다. • But 3 strand 이상의 것에 대해서는 새로운 experimental data가 필요할지도… • 우리의 model에 대해 (1)식을 적용하는 것이 타당할까? • quasi st-st의 적용은 타당할까?