Download
1 / 1
10 likes | 128 Views
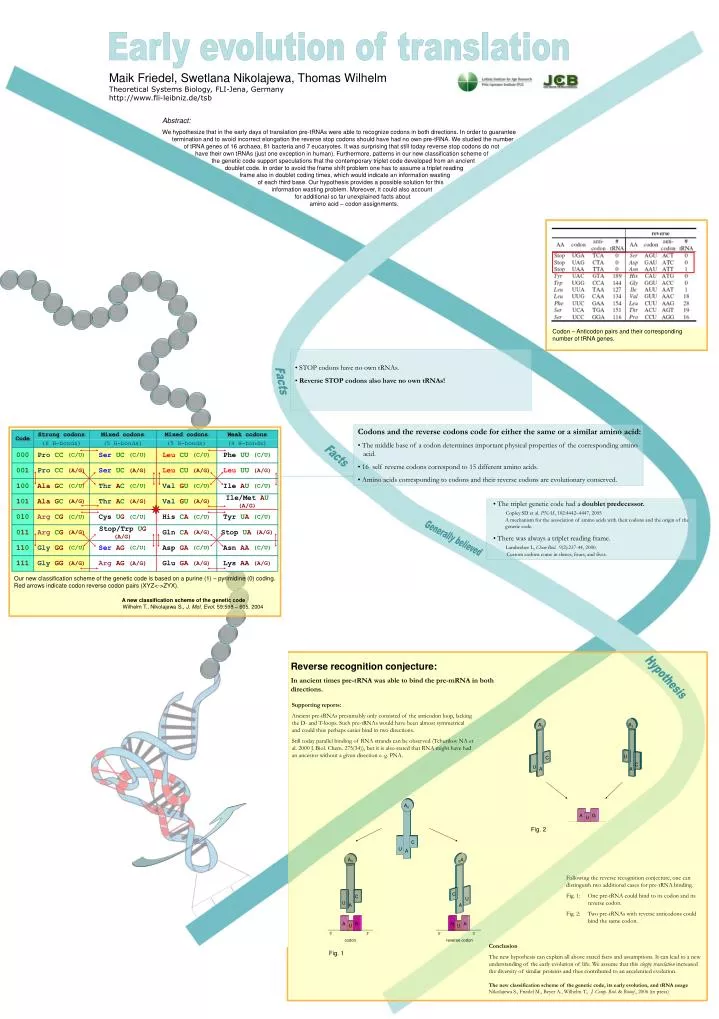
A 0. A. U. C. Code. Strong codons. Mixed codons. Mixed codons. Weak codons. (6 H-bonds). (5 H-bonds). (5 H-bonds). (4 H-bonds). 000. Pro CC (C/U). Ser UC (C/U). Leu CU (C/U). Phe UU (C/U). 00 1. Pro CC (A/G). Ser UC (A/G). Leu CU (A/G). Leu UU (A/G). 1 00.

E N D
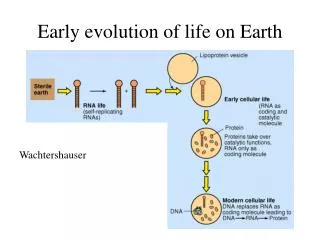
A0 A U C Code Strong codons Mixed codons Mixed codons Weak codons (6 H-bonds) (5 H-bonds) (5 H-bonds) (4 H-bonds) 000 Pro CC (C/U) Ser UC (C/U) Leu CU (C/U) Phe UU (C/U) 001 Pro CC (A/G) Ser UC (A/G) Leu CU (A/G) Leu UU (A/G) 100 AlaGC (C/U) ThrAC (C/U) Val GU (C/U) Ile AU (C/U) 101 AlaGC (A/G) Thr AC (A/G) ValGU (A/G) Ile/MetAU (A/G) 010 Arg CG (C/U) Cys UG(C/U) His CA (C/U) Tyr UA(C/U) 011 ArgCG (A/G) Stop/Trp UG (A/G) Gln CA(A/G) Stop UA (A/G) 110 Gly GG(C/U) Ser AG(C/U) Asp GA(C/U) Asn AA (C/U) 111 Gly GG (A/G) ArgAG(A/G) Glu GA(A/G) Lys AA (A/G) A0 A1 A A U C C U A G U A0 0A A A U C U C A N N A U U 5‘ 3‘ 5‘ 3‘ codon reverse codon Early evolution of translation Maik Friedel, Swetlana Nikolajewa, Thomas Wilhelm Theoretical Systems Biology, FLI-Jena, Germany http://www.fli-leibniz.de/tsb Abstract: We hypothesize that in the early days of translation pre-tRNAs were able to recognize codons in both directions. In order to guarantee termination and to avoid incorrect elongation the reverse stop codons should have had no own pre-tRNA. We studied the number of tRNA genes of 16 archaea, 81 bacteria and 7 eucaryotes. It was surprising that still today reverse stop codons do not have their own tRNAs (just one exception in human). Furthermore, patterns in our new classification scheme of the genetic code support speculations that the contemporary triplet code developed from an ancient doublet code. In order to avoid the frame shift problem one has to assume a triplet reading frame also in doublet coding times, which would indicate an information wasting of each third base. Our hypothesis provides a possible solution for this information wasting problem. Moreover, it could also account for additional so far unexplained facts about amino acid – codon assignments. Codon – Anticodon pairs and their corresponding number of tRNA genes. • STOP codons have no own tRNAs. • Reverse STOP codons also have no own tRNAs! Facts • Codons and the reverse codons code for either the same or a similar amino acid: • The middle base of a codon determines important physical properties of the corresponding amino acid. • 16 self reverse codons correspond to 15 different amino acids. • Amino acids corresponding to codons and their reverse codons are evolutionary conserved. Facts • The triplet genetic code had a doublet predecessor.Copley SD et al. PNAS, 102:4442–4447, 2005 A mechanism for the association of amino acids with their codons and the origin of the genetic code. • There was always a triplet reading frame. Landweber L, Chem Biol. 9(2):237-44, 2000. Custom codons come in threes, fours, and fives. Generally believed Our new classification scheme of the genetic code is based on a purine (1) – pyrimidine (0) coding. Red arrows indicate codon reverse codon pairs (XYZ<->ZYX). A new classification scheme of the genetic code Wilhelm T., Nikolajewa S., J. Mol. Evol. 59:598 – 605, 2004 Reverse recognition conjecture: In ancient times pre-tRNA was able to bind the pre-mRNA in both directions. Hypothesis Supporting reports: Ancient pre-tRNAs presumably only consisted of the anticodon loop, lacking the D- and T-loops. Such pre-tRNAs would have been almost symmetrical and could thus perhaps easier bind in two directions. Still today parallel binding of RNA strands can be observed (Tchurikov NA et al. 2000 J. Biol. Chem. 275(34)), but it is also stated that RNA might have had an ancestor without a given direction e. g. PNA. Fig. 2 Following the reverse recognition conjecture, one can distinguish two additional cases for pre-tRNA binding. Fig. 1: One pre-tRNA could bind to its codon and its reverse codon. Fig. 2: Two pre-tRNAs with reverse anticodons could bind the same codon. Conclusion The new hypothesis can explain all above stated facts and assumptions. It can lead to a new understanding of the early evolution of life. We assume that this sloppy translation increased the diversity of similar proteins and thus contributed to an accelerated evolution. The new classification scheme of the genetic code, its early evolution, and tRNA usage Nikolajewa S., Friedel M., Beyer A., Wilhelm T., J. Comp. Biol. & Bioinf., 2006 (in press) Fig. 1