Download

1 / 39
420 likes | 806 Views
Enzyme Properties and Kinetics. Andy Howard Introductory Biochemistry, Fall 2008 14 October 2008. Enzymes catalyze reactions. We need to classify them and get an idea of how they affect the rates of reactions. Classes of enzymes Enzyme kinetics Michaelis-Menten kinetics: overview

E N D
Enzyme Properties and Kinetics Andy HowardIntroductory Biochemistry, Fall 200814 October 2008 Biochemistry: Enzyme Properties
Enzymes catalyze reactions • We need to classify them and get an idea of how they affect the rates of reactions. Biochemistry: Enzyme Properties
Classes of enzymes Enzyme kinetics Michaelis-Menten kinetics: overview Kinetic Constants Kinetic Mechanisms Induced Fit Bisubstrate reactions Plans for Today Biochemistry: Enzyme Properties
Enzymes • Okay. Having reminded you that not all proteins are enzymes, we can now zero in on enzymes • Understanding a bit about enzymes makes it possible for us to characterize the kinetics of biochemical reactions and how they’re controlled Biochemistry: Enzyme Properties
Enzymes have 3 features • Catalytic power (they lower G‡) • Specificity • They prefer one substrate over others • Side reactions are minimized • Regulation • Can be sped up or slowed down by inhibitors and accelerators • Other control mechanisms exist Biochemistry: Enzyme Properties
IUBMB Major Enzyme Classes Biochemistry: Enzyme Properties
EC System Porcine pancreatic elastasePDB 3EST 1.65 Å 26kDa monomer • 4-component naming system,sort of like an internet address • Pancreatic elastase: • Category 3: hydrolases • Subcategory 3.4: hydrolases acting on peptide bonds (peptidases) • Sub-subcategory 3.4.21: Serine endopeptidases • Sub-sub-subcategory 3.4.21.36: Pancreatic elastase Biochemistry: Enzyme Properties
Category 1:Oxidoreductases • General reaction:Aox + Bred Ared + Box • One reactant often a cofactor (see ch.7) • Cofactors may be organic (NAD or FAD)or metal ions complexed to proteins • Typical reaction:H-X-OH + NAD+ X=O + NADH + H+ Biochemistry: Enzyme Properties
Category 2:Transferases • These catalyze transfers ofgroups like phosphate or amines. • Example: L-alanine + a-ketoglutarate pyruvate + L-glutamate • Kinases are transferases:they transfer a phosphate from ATP to something else Biochemistry: Enzyme Properties
O O- Category 3:hydrolases HO-P-O-P-OH O O- • Water is acceptor of transferred group • Ultrasimple: pyrophosphatase:Pyrophosphate + H2O ->2 Phosphate • Proteases,many other sub-categories Biochemistry: Enzyme Properties
C=C Category 4:Lyases • Non-hydrolytic, nonoxidative elimination (or addition) reactions • Addition across a double bond or reverse • Example: pyruvate carboxylase:pyruvate + H+ acetaldehyde + CO2 • More typical lyases add across C=C Biochemistry: Enzyme Properties
Category 5: Isomerases • Unimolecular interconversions(glucose-6-P fructose-6-P) • Reactions usually almost exactly isoergic • Subcategories: • Racemases: alter stereospecificity such that the product is the enantiomer of the substrate • Mutases: shift a single functional group from one carbon to another (phosphoglucomutase) Biochemistry: Enzyme Properties
Category 6: Ligases • Catalyze joining of 2 substrates,e.g.L-glutamate + ATP + NH4+L-glutamine + ADP + Pi • Require input of energy from XTP (X=A,G) • Usually called synthetases(not synthases, which are lyases, category 4) • Typically the hydrolyzed phosphate is not incorporated into the product Biochemistry: Enzyme Properties
iClicker quiz, question 1 • Collagenase catalyzes the cleavage of the -glycosidic bonds holding collagen together. Which IUBMB enzyme category would collagenase fall into? • (a) ligases (6) • (b) oxidoreductases (1) • (c ) hydrolases (3) • (d) isomerases (5) • (e) none of the above. Biochemistry: Enzyme Properties
iClicker quiz, question 2 • Triosephosphate isomerase, whose structure we discussed earlier, interconverts glyceraldehyde-3-phosphate and dhydroxyacetone phosphate. What would you expect the approximate DG value for this reaction to be? • (a) -30 kJ mol-1 (d) 0 kJ mol-1 • (b) 30 kJ mol-1 (e) no way to tell. • (c ) -14 kJ mol-1. Biochemistry: Enzyme Properties
Enzyme Kinetics • Kinetics: study of reaction rates and the ways that they depend on concentrations of substrates, products, inhibitors, catalysts, and other effectors. • Simple situation A B under influence of a catalyst C, at time t=0, [A] = A0, [B] = 0: • then the rate or velocity of the reaction is expressed as d[B]/dt. Biochemistry: Enzyme Properties
[B] Kinetics, continued t • In most situations more product will be produced per unit time if A0 is large than if it is small, and in fact the rate will be linear with the concentration at any given time: • d[B]/dt = v = k[A] • where v is the velocity of the reaction and k is a constant known as the forward rate constant. • Here, since [A] has units of concentration and d[B]/dt has units of concentration / time, the units of k will be those of inverse time, e.g. sec-1. Biochemistry: Enzyme Properties
More complex cases • More complicated than this if >1 reactant involved or if a catalyst whose concentration influences the production of species B is present. • If >1 reactant required for making B, then usually the reaction will be linear in the concentration of the scarcest reactant and nearly independent of the concentration of the more plentiful reactants. Biochemistry: Enzyme Properties
Bimolecular reaction • If in the reactionA + D Bthe initial concentrations of [A] and [D] are comparable, then the reaction rate will be linear in both [A] and [D]: • d[B]/dt = v = k[A][D] = k[A]1[D]1 • i.e. the reaction is first-order in both A and D, and it’s second-order overall Biochemistry: Enzyme Properties
Forward and backward • Rate of reverse reaction may not be the same as the rate at which the forward reaction occurs. If the forward reaction rate of reaction 1 is designated as k1, the backward rate typically designated as k-1. Biochemistry: Enzyme Properties
Multi-step reactions • In complex reactions, we may need to keep track of rates in the forward and reverse directions of multiple reactions. Thus in the conversion A B Cwe can write rate constantsk1, k-1, k2, and k-2as the rate constants associated with converting A to B, converting B to A, converting B to C, and converting C to B. Biochemistry: Enzyme Properties
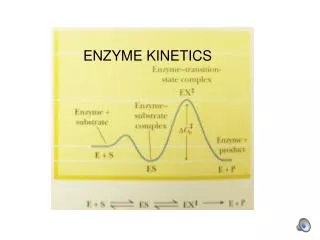
[ES] Michaelis-Menten kinetics t • A very common situation is one in which for some portion of the time in which a reaction is being monitored, the concentration of the enzyme-substrate complex is nearly constant. Thus in the general reaction • E + S ES E + P • where E is the enzyme, S is the substrate, ES is the enzyme-substrate complex (or "enzyme-intermediate complex"), and P is the product • We find that [ES] is nearly constant for a considerable stretch of time. Biochemistry: Enzyme Properties
Michaelis-Menten rates • Rate at which new ES molecules are being produced in the first forward reaction is equal to the rate at which ES molecules are being converted to (E and P) and (E and S). • Rate of formation of ES from left =vf = k1([E]tot - [ES])[S]because the enzyme that is already substrate-bound is unavailable! Biochemistry: Enzyme Properties
Equating the rates • Rate of disappearance of ES on right and left isvd = k-1[ES] + k2[ES] = (k-1+ k2)[ES] • This rate of disappearance should be equal to the rate of appearance • Under these conditions vf = vd. Biochemistry: Enzyme Properties
Derivation, continued • Thus since vf = vd. k1([E]tot - [ES])[S] = (k-1+ k2)[ES] • Km (k-1+ k2)/k1 = ([E]tot - [ES])[S] / [ES] • [ES] = [E]tot [S] / (Km + [S]) • But the rate-limiting reaction is the formation of product: v0 = k2[ES] • Thus v0 = k2[E]tot [S] / (Km + [S]) Biochemistry: Enzyme Properties
Maximum velocity • What conditions would produce the maximum velocity? • Answer: very high substrate concentration ([S] >> [E]tot),for which all the enzyme would be bound up with substrate. Thus under those conditions we getVmax = v0 = k2[ES] = k2[E]tot Biochemistry: Enzyme Properties
Using Vmax inM-M kinetics • Thus sinceVmax = k2[E]tot, • v0 = Vmax [S] / (Km+[S]) • That’s the famous Michaelis-Mentenequation Biochemistry: Enzyme Properties
Graphical interpretation Biochemistry: Enzyme Properties
Physical meaning of Km • As we can see from the plot, the velocity is half-maximal when [S] = Km • Trivially derivable: if [S] = Km, thenv0 = Vmax[S] / ([S]+[S]) = Vmax /2 • We can turn that around and say that the Km is defined as the concentration resulting in half-maximal velocity • Km is a property associated with binding of S to E, not a property of turnover Biochemistry: Enzyme Properties
kcat • A quantity we often want is the maximum velocity independent of how much enzyme we originally dumped in • That would be kcat =Vmax / [E]tot • Oh wait: that’s just the rate of our rate-limiting step, i.e. kcat = k2 Biochemistry: Enzyme Properties
Physical meaning of kcat • Describes turnover of substrate to product:Number of product molecules produced per sec per molecule of enzyme • More complex reactions may not have kcat = k2, but we can often approximate them that way anyway • Some enzymes very efficient:kcat > 106 s-1 Biochemistry: Enzyme Properties
Specificity constant, kcat/Km • kcat/Km measures affinity of enzyme for a specific substrate: we call it the specificity constant or the molecular activity for the enzyme for that particular substrate • Useful in comparing primary substrate to other substrates (e.g. ethanol vs. propanol in alcohol dehydrogenase) Biochemistry: Enzyme Properties
Kinetic Mechanisms • If a reaction involves >1 reactant or >1 product, there may be variations in kinetics that occur as a result of the order in which substrates are bound or products are released. • Examine eqns. 13.48, 13.49, 13.50, and the unnumbered eqn. on p. 430 in G&G, which depict bisubstrate reactions of various sorts. As you can see, the possibilities enumerated include sequential, random, and ping-pong mechanisms. Biochemistry: Enzyme Properties
Historical thought • Biochemists, 1935 - 1970 examined effect on reaction rates of changing [reactants] and [enzymes], and deducing the mechanistic realities from kinetic data. • In recent years other tools have become available for deriving the same information, including static and dynamic structural studies that provide us with slide-shows or even movies of reaction sequences. • But diagrams like these still help! Biochemistry: Enzyme Properties
Sequential, ordered reactions W.W.Cleland • Substrates, products must bind in specific order for reaction to complete A B P Q_____________________________E EA (EAB) (EPQ) EQ E Biochemistry: Enzyme Properties
Sequential, random reactions • Substrates can come in in either order, and products can be released in either order A B P Q EA EQ__E (EAB)(EPQ) E EB EP B A Q P Biochemistry: Enzyme Properties
Ping-pong mechanism • First substrate enters, is altered, is released, with change in enzyme • Then second substrate reacts with altered enzyme, is altered, is released • Enzyme restored to original state A P B QE EA FA F FB FQ E Biochemistry: Enzyme Properties
Induced fit Daniel Koshland • Conformations of enzymes don't change enormously when they bind substrates, but they do change to some extent. An instance where the changes are fairly substantial is the binding of substrates to kinases. Cartoon from textbookofbacteriology.net Biochemistry: Enzyme Properties
Kinase reactions • unwanted reactionATP + H-O-H ⇒ ADP + Pi • will compete with the desired reactionATP + R-O-H ⇒ ADP + R-O-P • Kinases minimize the likelihood of this unproductive activity by changing conformation upon binding substrate so that hydrolysis of ATP cannot occur until the binding happens. • Illustrates the importance of the order in which things happen in enzyme function Biochemistry: Enzyme Properties