Download
1 / 1
20 likes | 144 Views
Vertical Migration of Emerita talpoida in the Intertidal Zone Merete Ohme (PI), Philip Geyer, Julie Medina Department of Biological Sciences, Florida Institute of Technology. Background. Tidal Migrations.

E N D
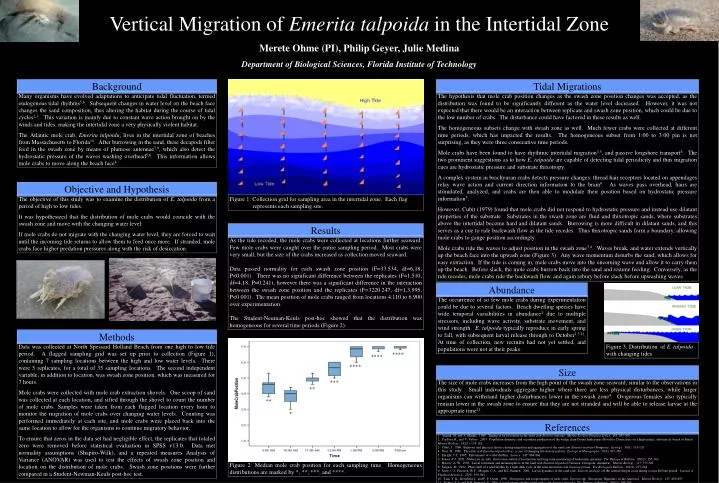
Vertical Migration of Emerita talpoida in the Intertidal Zone Merete Ohme (PI), Philip Geyer, Julie Medina Department of Biological Sciences, Florida Institute of Technology Background Tidal Migrations Many organisms have evolved adaptations to anticipate tidal fluctuation, termed endogenous tidal rhythms5,8. Subsequent changes in water level on the beach face changes the sand composition, thus altering the habitat during the course of tidal cycles2,3. This variation is mainly due to constant wave action brought on by the winds and tides, making the intertidal zone a very physically violent habitat. The Atlantic mole crab, Emerita talpoida, lives in the intertidal zone of beaches from Massachusetts to Florida10. After burrowing in the sand, these decapods filter feed in the swash zone by means of plumose antennae7,9, which also detect the hydrostatic pressure of the waves washing overhead5,6. This information allows mole crabs to move along the beach face4. The hypothesis that mole crab position changes as the swash zone position changes was accepted, as the distribution was found to be significantly different as the water level decreased. However, it was not expected that there would be an interaction between replicate and swash zone position, which could be due to the low number of crabs. The disturbance could have factored in these results as well. The homogeneous subsets change with swash zone as well. Much fewer crabs were collected at different time periods, which has impacted the results. The homogeneous subset from 1:00 to 3:00 pm is not surprising, as they were three consecutive time periods. Mole crabs have been found to have rhythmic intertidal migration3,5, and passive longshore transport4. The two prominent suggestions as to how E. talpoida are capable of detecting tidal periodicity and thus migration cues are hydrostatic pressure and substrate thixotropy. A complex system in brachyuran crabs detects pressure changes: thread hair receptors located on appendages relay wave action and current direction information to the brain6. As waves pass overhead, hairs are stimulated, analyzed, and crabs are then able to modulate their position based on hydrostatic pressure information5. However, Cubit (1979) found that mole crabs did not respond to hydrostatic pressure and instead use dilatant properties of the substrate. Substrates in the swash zone are fluid and thixotropic sands, where substrates above the intertidal become hard and dilatant sands. Burrowing is more difficult in dilatant sands, and this serves as a cue to ride backwash flow as the tide recedes. Thus thixotropic sands form a boundary, allowing mole crabs to gauge position accordingly. Mole crabs ride the waves to adjust position in the swash zone3,4. Waves break, and water extends vertically up the beach face into the upwash zone (Figure 3). Any wave momentum disturbs the sand, which allows for easy extraction. If the tide is coming in, mole crabs move into the oncoming wave and allow it to carry them up the beach. Before slack, the mole crabs burrow back into the sand and resume feeding. Conversely, as the tide recedes, mole crabs ride the backwash flow, and again rebury before slack before upwashing waves. Objective and Hypothesis The objective of this study was to examine the distribution of E. talpoida from a period of high to low tides. It was hypothesized that the distribution of mole crabs would coincide with the swash zone and move with the changing water level. If mole crabs do not migrate with the changing water level, they are forced to wait until the incoming tide returns to allow them to feed once more. If stranded, mole crabs face higher predation pressures along with the risk of desiccation. Figure 1: Collection grid for sampling area in the intertidal zone. Each flag represents each sampling site. Results As the tide receded, the mole crabs were collected at locations further seaward. Few mole crabs were caught over the entire sampling period. Most crabs were very small, but the size of the crabs increased as collection moved seaward. Data passed normality for each swash zone position (F=33.534, df=6,18, P<0.001). There was no significant difference between the replicates (F=1.510, df=4,18, P=0.241), however there was a significant difference in the interaction between the swash zone position and the replicates (F=3220.247, df=1,3.995, P<0.001). The mean position of mole crabs ranged from locations 4.110 to 6.900 over experimentation. The Student-Neuman-Keuls post-hoc showed that the distribution was homogeneous for several time periods (Figure 2). Abundance The occurrence of so few mole crabs during experimentation could be due to several factors. Beach dwelling species have wide temporal variabilities in abundance2 due to multiple stressors, including wave activity, substrate movement, and wind strength. E. talpoida typically reproduce in early spring to fall, with subsequent larval release through to October1,3,11. At time of collection, new recruits had not yet settled, and populations were not at their peaks. Methods Data was collected at North Spessard Holland Beach from one high to low tide period. A flagged sampling grid was set up prior to collection (Figure 1), containing 7 sampling locations between the high and low water levels. There were 5 replicates, for a total of 35 sampling locations. The second independent variable, in addition to location, was swash zone position, which was measured for 7 hours. Mole crabs were collected with mole crab extraction shovels. One scoop of sand was collected at each location, and sifted through the shovel to count the number of mole crabs. Samples were taken from each flagged location every hour to monitor the migration of mole crabs over changing water levels. Counting was performed immediately at each site, and mole crabs were placed back into the same location to allow for the organisms to continue migratory behavior. To ensure that zeros in the data set had negligible effect, the replicates that totaled zero were removed before statistical evaluation in SPSS v13.0. Data met normality assumptions (Shapiro-Wilk), and a repeated measures Analysis of Variance (ANOVAR) was used to test the effects of swash zone position and location on the distribution of mole crabs. Swash zone positions were further compared in a Student-Newman-Keuls post-hoc test. Figure 3: Distribution of E. talpoida with changing tides **** **** **** Size *** The size of mole crabs increases from the high point of the swash zone seaward, similar to the observations in this study. Small individuals aggregate higher where there are less physical disturbances, while larger organisms can withstand higher disturbances lower in the swash zone4. Ovigerous females also typically remain lower in the swash zone to ensure that they are not stranded and will be able to release larvae at the appropriate time11. ** ** * References 1. Amend, M. and A. Shanks. 1999. Timing of larval release in the mole crab Emerita talpoida. Marine Ecology Progress Series. 183: 295-300. 2. Cardoso, R., and V. Veloso. 2003. Population dynamics and secondary production of the wedge clam Donax hanleyanus (Bivalvia: Donacidae) on a high-energy, subtropical beach of Brazil. Marine Biology. 142(1): 153-162. 3. Cubit, J. 1969. Behavior and physical factors causing migration and aggregation of the sand crab Emerita analoga (Stimpson). Ecology. 50(1): 118-120. 4. Diaz, H. 1980. The mole crab Emerita talpoida (Say): a case of changing life history pattern. Ecological Monographs. 50(4): 437-456. 5. Enright, J.T. 1965. Entrainment of a tidal rhythm. Science. 147: 864-866. 6. Fraser, P.J. 2001. Statocysts in crabs: short-term control of locomotion and long-term monitoring of hydrostatic pressure. The Biological Bulletin. 200(2): 155-163. 7. Harvey, A. W. 1993. Larval settlement and metamorphosis in the sand crab Emerita talpoida (Crustacea: Decapoda: Anomura). Marine Biology. 117: 575-581. 8. Saigusa, M. 1992. Phase shift of a tidal rhythm by a light-dark cycle in the semi-terrestrial crab Sesarma pictum. The Biological Bulletin. 182(2): 257-264. 9. Sorte, C.J., Peterson, W.T. ,Morgan, C.A., and R.L. Emmett. 2001. Larval dynamics of the sand crab, Emerita analoga, off the central Oregon coast during a stron El Nino period. Journal of Plankton Research. 23(9): 939-941. 10. Tam, Y. K., Kornfield, I., and F. P. Ojeda. 1996. Divergence and zoogeography of mole crabs, Emerita spp. (Decapoda: Hippidae), in the Americas. Marine Biology. 125: 489-497. 11. Ziegler, T.A. and R.B. Forward, Jr. 2005. Larval release rhythm of the mole crab Emerita talpoida. The Biological Bulletin. 209(3): 194-203. Figure 2: Median mole crab position for each sampling time. Homogeneous distributions are marked by *, **, ***, and ****.






















