Download

1 / 58
590 likes | 791 Views
IV. Přenos signálů-buněčné receptory. Úvod Sedm transmembránových helixových receptorů mění svoji konformaci po vazbě ligandu za současné aktivace G proteinů Hydrolýza fosfatidylionositolbisfosfátů fosfolipasou C vede ke tvorbě dvou poslů Vápenatý iont jako univerzální cytosolární posel

E N D
IV. Přenos signálů-buněčné receptory • Úvod • Sedm transmembránových helixových receptorů mění svoji konformaci po vazbě ligandu za současné aktivace G proteinů • Hydrolýza fosfatidylionositolbisfosfátů fosfolipasou C vede ke tvorbě dvou poslů • Vápenatý iont jako univerzální cytosolární posel • Dimerizace receptorů jako odezva na vazbu ligandu • Důsledky defektů přenosu signálů
Formy transdukce signálu • Receptory na membráně přenáší informaci z okolí do buňky. Některé nepolární signální molekuly (např. steroidní hormony) pronikají do buněk a tam se váží na protein nebo přímo na DNA. • Častější interkace ligand – receptor. Receptor je obvykle integrální protein membrány. Uvnitř buňky se tvoří druhý posel. Ireversibilní. • Obecnou cestou přenosu informace je fosforylace - kinasy. Zasaženými místy na enzymech jsou Ser, Thr a Tyr. Reversibilní. Terminace přenosu informace – proteinfosfatasy.
Sedm transmebránových helixů – 7TM • Receptory tohoto typu přenáší signály fotonů, odorantů, chutí, hormonů a neurotransmiterů.
Příkladem hormonu, který se váže na sedm transmembránový receptor je adrenalin.
Biologické funkce zprostředkované receptory 7TM Vůně Chuť Vidění Přenos nervových vzruchů Uvolňování hormonů Chemotaxe – je pohyb vyvolaný chemikáliemi Exocytosa - proces zajišťující vylučování látek z buněk do extracelulárního prostoru. Kontrola tlaku krve Embryogeneze – vývoj zárodku (embrya) Růst a diferenciace buněk Virální infekce Karcinogeneze
Rhodopsin – (opsin + retinal) a navazující heterotrimerní G-protein.
Konformační změny Ga po výměně nukleotidů. Aby mohlo dojít k výměně nukleotidů dochází vlivem vstupu signálu na receptor přes 7TM ke třem konformačním změnám (změna, přepnutí).
Conceptual insights – Signál – přenos • E:\MREG2010\Biochemistry, 6e - Content.mht
Rodiny G-proteinů (heterotrimerů) a jejich funkce Gasb-Adrenergní aminy, Stimulují glukagon, parathyreoidní hormon, adenylátcyklasu mnoho dalších Gai Acetylcholin, a-adrenergní aminy, Inhibují mnoho neurotransmiterů adenylátcyklasu Gat Fotony Stimulují cGMP fosfodiesterasy Ga13 Thrombin, další agonisté Stimulují Na+ a H+ výměnu Gaq Acetylcholin, a-adrenergní aminy, Zvyšují IP3 a mnohé neurotransmitery intracelulární [Ca2+]
Adrenergní • Adrenergní – nerv, buňka nebo buněčný receptor (adrenoreceptor) aktivovaný adrenalinem nebo noradrenalinem a analogickými látkami. • Všechny adrenergní receptory se běžně dělí na • a-adrenoreceptory a b-adrenoreceptory • Receptory skupiny a jsou citlivější k adrenalinu než k izoproterenolu, b-adrenoreceptory reagují přesně naopak. V těle se v typickém případě váže noradrenalin spíše na a-adrenoreceptory, zatímco adrenalin na adrenoreceptory skupiny b; existuje však mnoho výjimek. Rozlišují se ještě a1 a a2 adrenoreceptory, stejně tak β1 a β2 adrenoreceptory. • Funkce adrenergních receptorů jsou velmi různé a liší se podle konkrétního typu adrenergního receptoru. • a-Adrenoreceptory: zvýšená glykogenolýza v játrech , zvýšená glukoneogeneze, relaxace hladkého svalstva ve střevech. • b-Adrenoreceptory : zvýšená svalová glykogenolýza a jaterní glukoneogeneze a glykogenolýza, mobilizace zásobního tuku, zrychlení srdeční frekvence a prohloubení srdečních stahů.
Isoprotenerol, isoprenalin (vlevo), adrenalin (vpravo). Sympatomimetikum betaadrenergní agonista – shodný efekt s adrenalinem. Využívá se při zástavách srdce, astmatických záchvatech apod. β1 agonist: stimuluje aktivitu adenylátcyklasy; otevírá Ca2+ kanálky. (srdeční stimulátor; používá se při akutním kolapsu srdce a arytmiích)
Terminace signálu - obnova Ga. GTPasová aktivita hydrolyzuje GTP a Ga se znovu spojí s podjednotkami b a g.
Terminace signálu (např. adrenalin). A) Disociace signální molekuly z receptoru. B) Fosforylace cytoplasmatického C terminálního konce receptoru GRK kinasoua následná vazba b-arrestinu.
Arrestin • Vypnutí signálu arrestinem. • Pevním krokem je fosforylace kinasou z řady Ser/Thr kinas zvanou G protein Coupled Receptor Kinase (GRK). • GRK fosforylace specificky připraví aktivovaný receptor pro vazbu arrestinu. • Arrestin vázaný na receptor brání dalšímu přenosu signálu cestou G proteinu. • Arrestin se také váže na jiné třídy receptorů na memebráně a na další proteiny.
Krystalografická struktura hovězího arrestinu – S. S-antigen; sítnicový.
Aktivace adenylátcykalsy Gas.A)Adenylátcyklasa je integrální membránový (12 helixů) protein. Katalyticky aktivní jsou dvě intracelulární domény. B) Gas s navázaným GTP se váže na katalytickou část cyklasy a stimuluje její aktivaci.
Cyklický AMP • V eukaryotních buňkách aktivuje proteinkinasu A (PKA). • PKA je složena ze dvou regulačních a dvou katalytických řetězců. Za nepřítomnosti cAMP je komplex katalyticky inaktivní. Po vazbě cAMP na regulační řetězce je PAK aktivována – katalyzuje fosforylaci specifických Ser a Thr proteinů, které jsou takto aktivovány. • Např. aktivace štěpení glykogenu (glykogenfosforylasa a-aktivni a glykogenfosforylasa b - inaktivni) a současně inhibuje syntézu glykogenu.
Inhibitory fosfodiesterasy • Purinové alkaloidy – kofein, theobromin, theofylin. • Vyskytují se v kávě a čaji.
Fosfolipasy – A1,,A2, C a D. Působení fosfolipas A1,aA2 = fosfolipasa B. Phospholipase C - štěpí před fosfátem. Uvolňuje diacyglycerol a skupinu obsahující fosfátovou hlavičku. Fosfolipasy C hrají centrální roli při přenosu signálu tím, že uvolňují druhého posla – inositoltrifosfát.
Hydrolýza fosfatidylinositolbisfosfátu (membránový lipid) fosfolipasou C vede ke tvorbě dvou druhých poslů. Např. po vazbě vasopressinu (antidiuretický hormon, ADH) na 7TM. Dva poslové: inositol-1,4,5-trifosfát (IP3) a diacylglycerol
Cys-Tyr-Phe-Gln-Asn-Cys-Pro-Arg-Gly-NH2 N-konec C-konec; nonapeptid Syntetizován v hypothalmu jako preprohormon, skladován ve váčcích zadního laloku.
Oxytocin - nonapeptid • Cys-Tyr-Ile-Gln-Asn-Cys-Pro-Leu-Gly-NH2 • Oxytocin jepeptidový hormon savců, který se syntetizuje v hypothalmickém jádrunucleusparaventricularis. • Dokrevního oběhu je uvolňován z neurohypofýzy Uvolňuje se u žen po stimulaci prsu, dělohy a vagíny.
Inositol 1,4,5-trisfosfát otevírá kanály a uvolňuje tak Ca2+ z intracelulárních prostor. • Působením IP3 dochází k uvolnění Ca2+ z endoplasmatického retikula, buněk hladkého svalstva a sarkoplasmatického retikula. • Zvýšená hladina Ca2+ v cytosolu vede např. ke kontrakci hladkého svalstva a štěpení glykogenu. • Jak se signál IP3 vypíná ? • IP3 se velmi rychle přeměňuje na další deriváty inositolfosfátů a na inositol (fosfatasy). Jeho délka života ve většině buněk je několik sekund. • Recyklaci IP3 inhibuje Li+ !!!
Metabolismus IP3: Tvorba a odbourávání fosfátových derivátů inositolu.
Diacylglycerol aktivuje proteinkinasu C (PKC) – ta fosforyluje mnoho cílových proteinů.
Metabolismus diacylglycerolu. Diacylglycerol je buď fosforylován na fosfatidát nebo hydrolyzován na glycerol a mastné kyseliny.
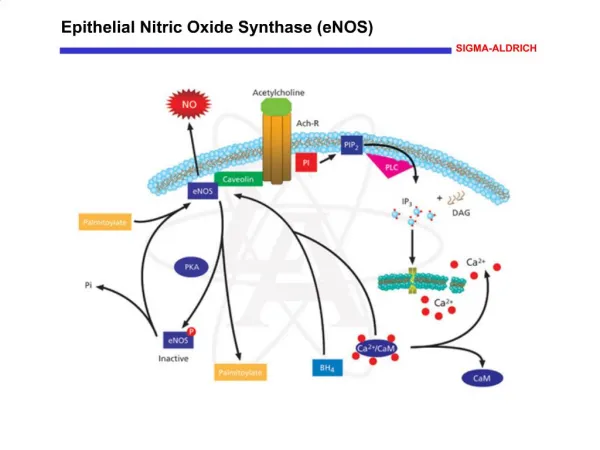
Ca2+ se specificky váže na calmodulin – malý kyselý protein o zhruba 148 AMK, (16 706D). Obsahu EF motiv !! Calmodulin (CaM) (zkratka CALciumMODULatedproteIN)
CaM se účastní procesů jako je zánět, apoptosa, stahy hladkého svalstva, vnitrobuněčný pohyb, krátko- i dlouhodobá paměť, imunitní odpověď a růst nervové tkáně. CaM jako součást kinasy.
Vápenatý iont je univerzální cytosolární posel. Na obr. je detail vazby Ca2+ na kalmodulin.
Proč je Ca2+ univerzálním druhým poslem ? • 1. Vápenatý iont vytváří komplexy s řadou buněčných složek. Komplexy jsou nerozpustné. Proto je nutné udržovat hladinu Ca2+ na nízké hladině. To zajišťují transportní kanálové systémy. • U eukaryot jsou nejdůležitější Ca2+ ATPasa a Na+ - Ca2+ výměny. • Díky těmto pumpámje cytosolární koncentrace Ca2+ udržována u neexcitovaných buněk na 100 nM !! To je několika řádově nižší koncentrace ve srovnání s krví (5 mM). • 2. Vápenaté ionty se váží pevně na proteiny. Vazba na vedlejší řetězce Glu a Asp a kyslík bez náboje (karbonyly řetězce a kyslíkové atomy Asn a Gln. • Jakým způsobem lze měřit koncentraci Ca2+ v buňkách ??
Fluorescenční spektrum barviva vážícího vápenaté ionty (Fura-2). Měří se tak koncentrace v buňkách i v prostředí.
Vápenaté ionty aktivují regulační protein calmodulin, který stimuluje mnoho enzym a přenašečů. • Calmodulin je 17 kD protein (CaM), obsahujícíčtyři vazebná místa pro Ca2+. • Calmodulin je aktivován, když se intracelulární hladina Ca2+ zvedne nad 500 nM !!! • Calmodulin patří do rodiny EF-ruky proteinů. EF-ruka je vazebný motiv Ca2+. • Calmodulin s navázanými Ca2+aktivuje řadu enzymů, iontových pump a dalších cílových proteinů. • Ca2+/calmodulin-dependent protein kinases neboli CaM kinases (EC2.7.11.17) jsou serin/threoninové prroteinkinasy primárně regulované komplexem Ca2+/calmodulin.
Druhá skupina proteinů vážících Ca2+. • Proteiny vážící vápenaté ionty se nazývají annexiny – váží vápenaté ionty a fosfolipidy. • Do roku 2002 bylo identifikováno 160 annexinových proteinů v 65 druzích. • Kritérium pro annexiny: Váží fosfolipidy s negativním nábojem v prostředí vápenatých iontů a obsahují 70 aminokyselinovou opakující se sekvenci (annexinové opakování).
Dimerizace receptorů jako odezva na vstup ligandu a přenos signálu křížovou fosforylací. • Jako příklad uvedeme lidský růstový hormon. Monomerní protein, 217 aminokyselin, čtyři helixy. • Receptor je protein 638 aminokyselin – dělí se na část extracelulární (250 aminokyselin), jednoduchý helix přes membránu a intracelulární doména (350).
Křížová fosforylace JAKů (proteinkinasa, Janusova kinasa 2). Obě fosforylasy se navzájem fosforylují a tím se aktivují. Janusova kinasa (JAK = Just Another Kinase) je cytoplasmatická tyrosinkinasa.