Download

1 / 47
510 likes | 748 Views
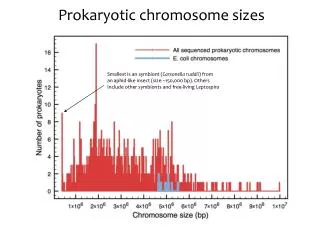
Gene Expression Prokaryotic Gene Transcription (Cont.) & Eukaryotic Transcription Histones and Chromatin G & G Pages 336-340, Chapter 29, 9/14/11 Thomas Ryan, Ph.D. Biochemistry and Molecular Genetics tryan@uab.edu. Prokaryotic Chromosome ( E. coli ). Large circular chromosome - 4.6 x 10 6 bp

E N D
Gene ExpressionProkaryotic Gene Transcription(Cont.)&Eukaryotic TranscriptionHistones and ChromatinG & G Pages 336-340, Chapter 29, 9/14/11Thomas Ryan, Ph.D. Biochemistry and Molecular Geneticstryan@uab.edu
Prokaryotic Chromosome (E. coli) • Large circular chromosome - 4.6 x 106 bp • Genome forms a compact structure called the nucleoid • DNA organized in 50-100 loops (domains) • The ends of loops are constrained by binding to protein structure which is in contact with cell membrane
General Organization of Operons Operators can be upstream, downstream, or overlapping with the promoter. Regulatory proteins that bind to the operator can influence the access of RNA polymerase to the promoter thereby affecting the rate of transcription initiation.
Induction and Repression • Increased synthesis of genes in response to metabolites is ‘induction’ and the metabolites are called co-inducers • Decreased synthesis in response to a metabolite is termed ‘repression’ and the metabolite a co-repressor • Some substrates induce enzyme synthesis even though the enzymes can’t metabolize the substrate - these are ‘gratuitous inducers’ - such as IPTG
lac OperonBoth Positive and Negative Regulation (CRP•cAMP)2 binding RNA Polymerase binding -61.5 -35 -10 Repressor binding +1 mRNA Multiple control mechanisms are the norm.
The lac OperonNegative regulation • The structural genes of the lac operon are controlled by negative regulation • lacI gene product is the lac repressor • lacI mutants express the genes needed for lactose metabolism constitutively • The lac operator has palindromic DNA • lac repressor - forms tetramer; DNA binding in the N-terminal domain; Inducer binding at the C-terminal domain
lac Operon • Negative Regulation • Operon is generally “off”; only fully “on” when lactose is present and glucose is absent • When no lactose is present: repressor is bound; inhibiting transcription elongation through the operon • When lactose is present: lactose is converted to allolactose, binds to repressor, causing it to fall off the operator and allowing transcription of the downstream cistrons to proceed
Gratuitous Inducers: Not substrates, but still induce -Galactoside linkage CH3 CH2OH CH2OH OH HO HO S C H O O O OH OH OH CH3 OH O OH OH CH2OH Isopropyl -D-thiogalactoside Lactose (IPTG) substrate not a substrate
-galactosidase (LacZ) CH2OH OH O OH glucose HO H2O CH2OH OH OH HO O O OH OH CH2OH OH O HO OH -gal -gal O OH CH2OH OH -gal -gal lactose galactose OH Other enzymes in the lac system lactose permease (LacY) acetylase (LacA)
lac Operator Sequence • The operator site is palindromic and lies just downstream of the transcription initiation site • lac repressor is tetrameric and blocks transcription elongation by RNAP
Induction of the lac Operon Binding of -galactosides to the lac repressor induces a cooperative allosteric change. The inducer:lac repressor complex dissociates from the operator and transcription of the structural genes occurs.
lac Operon: Catabolite Repression E. coli can use several sugars as carbon sources but prefers glucose because requires no energy to take it up Glucose-sensitive operons: their expression is reduced in the presence of glucose Catabolism of glucose inhibits expression of these operons – called catabolite repression This mechanism allows for the lac genes to be only partially induced in the presence of lactose and glucose
Catabolite Activator ProteinPositive Control of the lac Operon • Some promoters require an accessory protein to speed transcription • Catabolite Activator Protein (CAP) is one such protein • CAP is a homodimer of 22.5 kD peptides • N-term binds cAMP; C-term binds DNA • Binding of CAP-(cAMP)2 to DNA assists formation of closed promoter complex [also called CRP (cAMP receptor protein)]
Adenine O CH2 O P O OH O O 3, 5 cyclic AMP (cAMP) Catabolite Activator Protein Mechanism of Activation Glucose starvation activates adenylyl cyclase increasing cAMP levels. cAMP acts as a co-inducer binding to CAP and activating transcription by binding to upstream promoter sequences.
= cAMP Catabolite Activator Protein Upstream promoter sequence Binding of CAP to CAP site (e.g. E. coli lac operon) induces a bend in the DNA
The trp OperonCo-repressor Mediated Negative Control • Encodes a leader sequence and 5 enzymes for tryptophan biosynthesis • trp operon is always “on” unless tryptophan levels are sufficiently high to turn “off” • Tryptophan is the co-repressor • Trp repressor binding excludes RNA polymerase from the promoter • Trp repressor also regulates trpR and aroH operons and is itself encoded by the trpR operon. This is autogenous regulation (autoregulation) - regulation of gene expression by the product of the gene.
trp Operon Five structural genes transcribed from the single promoter repressor
Regulation of the trp Operon by Repression and Attenuation Inactive Active
Attenuation • The need for tryptophan is tested during translation of the “leader” sequence (“trpL”) on all mRNA initiated at promoter. • trp expression is modulated by the tRNA-trp (“charged tRNA”) available to translate codon UGG. If the ribosomes translate these codons rapidly, transcription will stop because: • Different segments in the leader mRNA sequence can pair with each other to form alternative stem-loop structures. • If ribosomes “stall” on the trp codons, the RNA structure formed is not a terminator. TRANSCRIPTION OF trp OPERON PROCEEDS • If ribosomes translate the leader and pass the trp codons quickly, an intrinsic terminator is formed. TRANSCRIPTION OF trp OPERON TERMINATES
“Pause Hairpin” “Antiterminator” “Terminator”
Gene ExpressionEukaryotic Gene Transcription Histones and Chromatin
Eukaryotic Vs. Prokaryotic Transcription • Presence of a nucleus means transcription and translation occur in separate compartments of the cell • Larger genomes (1000X between humans and E. coli) • Chromatin structure in eukaryotes limits accessibility • Three RNA polymerases • Eukaryote pre-mRNAs are subject to extensive post-transcriptional modification
Genome Sizes of Various Organisms The human genome contains approximately 1000 times the amount of DNA compared to E. coli. However, humans have only 20 times as many genes as E. coli. (98.5% of the human genome is noncoding compare to only 11% of the E. coli genome).
Eukaryotic Chromatin Structure • The human genome contains 3 x 109 bp per haploid genome. • If the DNA of all 46 chromosomes from one cell was linked together, it would measure one meter in length. • If the DNA in all cells of a single human was linked end to end, it would stretch to the sun and back. • How is the DNA packaged into the nucleus such that genes are still accessible to the transcriptional regulatory proteins that control their expression?
LINES - Long interspersed nuclear elements (6.1 kb full length) SINES - Short interspersed nuclear elements (300 bp) Characterization of Human Genomic DNA
Eukaryotic Chromatin Structure • Each eukaryotic chromosome contains a single linear supercoiled DNA molecule • Nucleoprotein material of the eukaryotic chromosome is called chromatin(complex of DNA, protein and RNA)
Nuclear membrane Nuclear pore Heterochromatin Cytoplasm Nucleus Euchromatin EM of a liver cell: cytoplasm (left) separated from the nucleus (right) by the nuclear membrane (middle)
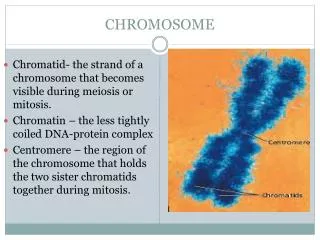
Eukaryotic Chromosome Structure • Euchromatin- comprises most of the genome, transcriptionally active, susceptible to DNaseI digestion • Heterochromatin- highly condensed inactive chromatin located at centromeres and telomeres (less susceptible to DNase digestion) • Centromere- attachment point for sister chromatids and spindle fibers • Telomere- ends of chromosome
Eukaryotic Chromatin Structure • DNA associates with histones proteins to produce nucleosomes • Nucleosome- the fundamental unit of organization of the chromatin fiber • Each nucleosome contains a core particleof basic proteins – histones - which aresurrounded by DNA • Electron microscopy shows fiber structure of chromosomes as “beads on a string”
Histones • Small basic proteins, rich in lysine and arginine • Interact with DNA through electrostatic interactions • Five major types of histones: • H1, H2A, H2B, H3 and H4 • H2A, H2B, H3 and H4 form a complex of 8 proteins • DNA is supercoiled around histone octets forming nucleosomes • H1 associates with neighboring nucleosomes to form a more closely packed structure 30nm solenoid
Histone Structure (B) Structure of the histone fold. (C) Histones 2A and 2B form a dimer through an interaction known as the handshake. Histones H3 and H4 form a dimer through the same type of interaction.
Histone Structure • H2A and H2B monomers form a H2A-H2B dimer. • Similarly, H3 and H4 monomers form an H3-H4 dimer.
Histone Structure • H2A-H2B and H3-H4 dimers bind to form tetramers. • Two tetramers will come together to form the 8 histone octet
DNA Wraps Around Histone Octomers • The DNA helix makes 1.65 turns around the histone octamer. • 146 base pairs of DNA wraps around each histone octet. • Histones are among the most highly conserved eukaryotic proteins. For example, the amino acid sequence of histone H4 from a pea and a cow differ at only 2 of the 102 amino acids.
Solenoid Formation: Role of H1 Histone Formation of the 30nm fiber requires Histone H1 which is a globular histone that is highly conserved. Histones are positively charged and this helps to compact the negatively charged DNA.
Histone Tails • Nucleosomal Core Without DNA • Yellow: H2A • Red: H2B • Blue: H3 • Green: H4 • Note ordered structure of histone amino terminal tails. These tails can contact neigh-boring nucleosomes. • When the tails are acetylated, interactions between nucleo-somes are diminished and chromatin becomes more extended or open.
Eukaryotic Chromatin Structure Electron micrographs of 30nm “Solenoid” structure (top) and 10nm DNA/histone “beads on a string” structure (bottom).
Eukaryotic Chromatin Structure Looping and Miniband
Eukaryotic Chromatin Structure • Chromosomes • attachment of loops of chromatin fibers to nonhistone protein complexes =scaffold • Nucleosome fibers condense more into 30 nm chromatin fiber. Predominant form in interphase nucleus • Duplex DNA winds around histone octamers to form nucleosomes= 11nmhistone fiber • Primary structure of DNA = double helix = 2nm duplex DNA