Download

1 / 37
470 likes | 1.05k Views
Principles that Govern Protein Structure and Stability. Interactions between atoms within the protein chain Interactions between the protein and the solvent . Forces that stabilize protein structure. Covalent bonds Hydrogen bonds Metal ligands Ionic interactions Disulfide bonds

E N D
Interactions between atoms within the protein chain Interactions between the protein and the solvent Forces that stabilize protein structure
Covalent bonds Hydrogen bonds Metal ligands Ionic interactions Disulfide bonds Non-bond interactions Bond types in proteins
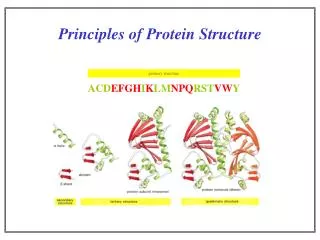
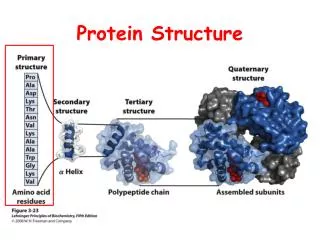
Covalent interactions establish the structural framework of the protein molecule, the chemical expression of primary structure Backbone conformation constrained by steric restrictions on and torsions Sidechain conformations are also constrained Favourable sidechain conformations depend on the sidechain and also on its neighbours. S-S Bonds between cysteine residues can form within proteins Favourable conformations in polypeptides
Charged side chains in protein can interact favorably with an opposing charge of another side chain according to Coulomb’s law: Examples of favorable electrostatic interaction include that between positively charged lysine and negatively charged glutamic acid. Salts have the ability to shield electrostatic interactions. Electrostatic Interactions
Coulomb interaction between two ions At close range, Coulomb interactions are as strong as covalent bonds Their energy decreases with 1/r and fall off to less than kT at about 56 nm separation between charges In practice, charge-charge interactions have been shown to be chemically significant at up to 15 Å in proteins Small charged metal ions can act as positive charge in an ion pair Charge-charge interactions Salt Bridge Mg-ATP
Noncovalent chemical bond in which an electronegative atom (a hydrogen-bond acceptor) shares a hydrogen atom with an electronegative atom with a bound hydrogen Energy : 10-40 kJ/mol Approximately 1.7-3 Å in length Strength varies with angle of hydrogen-bond interaction Individually, not very strong, but the large numbers of hydrogen bonds in regular secondary structures stabilize the framework of the protein Hydrogen bonds O H N C
van der Waals forces • van der Waals forces are also known as London forces. • They are weak interactions caused by momentary changes in electron density in a molecule. • They are the only attractive forces present in nonpolar compounds. Even though CH4 has no net dipole, at any one instant its electron density may not be completely symmetrical, resulting in a temporary dipole. This can induce a temporary dipole in another molecule. The weak interaction of these temporary dipoles constituents van der Waals forces. The surface area of a molecule determines the strength of the van der Waals interactions between molecules. The larger the surface area, the larger the attractive force between two molecules, and the stronger the intermolecular forces.
Dispersion A neutral atom: argon. It is like a large spherical jelly with a golf ball at the centre. The golf ball is the nucleus carrying a large positive charge and the jelly represents the clouds of electrons. At a point external to the atom the net average field will be zero because the positively-charged nucleus' field will be exactly balanced by the electron clouds. However, atoms vibrate (even at 0K) and so that at any instant the cloud is likely to be slightly off centre. This disparity creates an "instantaneous dipole":
Dispersion Suppose that we have another argon atom close to the first. This atom will see the electric field resulting from the instantaneous dipole. This field will effect the the electron and induce a dipole The two dipoles attract one another - producing an attractive interaction. The Dispersion interaction can be shown to vary according to the inverse sixth power of the distance between the two atoms: B depends on the polarizability of the atoms r is the distance between them
Repulsion When two atoms are brought increasing close together there is a large energetic cost as the orbitals start to overlap. The Pauli exclusion principle states that no two electrons can share the same state so that in effect half the electrons of the system would have to go into orbitals with an energy higher than the valence state. For this reason the repulsive core is sometimes termed a "Pauli exclusion interaction". The Hard Sphere Model: atoms have a characteristic radius (below the van der Waals radius) and cannot overlap. Can represent the energy costs of close approach using a term
Nonspecific forces between like or unlike atoms Decrease with r6 approximately 1 kJ/mol If r0 is the sum of van der Waals radii for the two atoms. Van der Waals forces are attractive forces when r> r0 and repulsive when r< r0. van der Waals forces
The hydrophobic force • Observation: • Hydrophobic residues are buried while hydrophilic residues are on the outside. • The exterior surface area of proteins can be up to 60% polar atoms • Proteins fold to maximize their effectiveness as hydrogen-bonding partners to water • Explanation: • When hydrophobic residues are exposed to solvent, the extended hydrogen bonding structure of water is disrupted • Breaking hydrogen bonds in water is energetically unfavourable • Water molecules reorient around the hydrophobic molecule, so that the least number of hydrogen bonds are sacrificed to accommodate it • Burying hydrophobic residues releases water and increases entropy.
Protein Interior and Exterior Internal packing of atoms in a protein can be analysed by depicting every atom in the protein as a sphere with the appropriate van der Waals radius. Overlapping regions (regions of covalent bonds) are truncated. This surface is called the van der Waals surface. Cannot measure a van der Waals surface of a protein because any chemical probe will have some dimension greater than zero. A more realistic representation is the solvent accessible surface that is defined by the center of a water molecule (sphere with radius 1.4 Å) as it moves over the surface of a protein.
Protein Interior and Exterior A more realistic representation is the solvent accessible surface that is defined by the center of a water molecule (sphere with radius 1.4 Å) as it moves over the surface of a protein. Protein-protein interactions, which form the basis for most cellular processes, result in the formation of protein interfaces. The protein-protein docking problem is the prediction of a complex between two proteins given the three-dimensional structures of the individual proteins
Secondary structures pack closely to one another and also intercalate with extended polypeptide chains Most polar residues face the outside of protein and interact with solvent but may be buried if H-bonding and charge is satisfied Most hydrophobic residues face the interior of the protein and interact with each other thereby minimizing contact with water van der Waal’s volume is about 72-77% of the total protein volume; about 25% is not occupied by protein atoms. These cavities provide flexibility in protein conformation and dynamics Random coil or loops maybe of importance in protein function (interacting with other molecules, enzyme reactions) Packing of Globular Proteins
Folding of a globular protein is a thermodynamically favored process, i.e. G must be negative. The folding process involves going from a multitude of random-coil conformations to a single folded structure. The folding process involves a decrease in randomness and thus a decrease in entropy -S and an overall positive contribution to G. This decrease in entropy is termed “conformational entropy”. An overall negative G : a result of features that yield a large negative H or some other increase in entropy on folding. The Thermodynamics of Folding G = H - TS
Formation of secondary structure is an enthalpy driven process Energy derived from the formation of many van der waals and H-bonding interactions as well as the alignment of dipoles overcomes the loss of entropy associated with the formation of the peptide backbone conformation. Formation of tertiary structure is enthalpically unfavorable Energy loss in the burying of ion-pairs (~1 kcal/mol) and the breaking of shorter, stronger H2O bonds. Though some energy is gained from van der waals packing, very little is gained from the formation of internal h-bonds because as many h-bonds with water are broken as are formed in the process of folding a protein. Free energy associated with solvation of an ion is ~-60 kcal/mol An ion will NOT be buried in the hydrophobic interior of a protein. Protein Folding : No Net Enthalpic Contribution
Upon protein folding hydrophobic residues move to the interior of the protein caged H2O molecules are released Enthalpy is gained : unfavorable (H +) entropy is also gained (S +) : extremely favorable Increase in entropy of water compensates for the loss of conformational entropy of the protein and drives the protein folding process Protein Folding : Entropy Driven Process
Difference in energy (free energy) between folded (native) and unfolded (denatured) state is small, 5-15 kcal/mol Enthalpy and entropy differences balance each other, and DG is a small positive number. Small DG is necessary because too large a free energy change would mean a very stable protein, one that would never change However, structural flexibility is important to protein function, and proteins need to be degraded Free energy of folding
Random Coil Native conformation Protein Folding • What are the forces that guide this process? • What are the Steps Involved? • How Fast Can this Happen?
The Thermodynamic Hypothesis “The native, folded structure of a protein, under optimal conditions, is the most energetically stable conformation possible” Christian Anfinsen, 1972 • Most of the information for determining the three-dimensional structure of a protein is carried in its amino acid sequence Anfinsen, C.B. Principles that govern the folding of protein chains. Science181, 223-30 (1973).
Consider a protein of 100 amino acids. Assign 2 conformations to each amino acid. The total conformations of the protein is 2100=1.27x1030. Allow 10-13 sec for the protein to sample through one conformation in search for the overall energy minimum. The time it needs to sample through all conformations is (10-13)(1.27x1030)=1.27x1017sec = 4x109 years! Levinthal’s paradox illustrates that proteins must only sample through limited conformations, or fold by “specific pathways”. Much research efforts are devoted in searching for the principles of the “specific pathways”. Levinthal’s Paradox
For any given protein, there is one conformation that has significantly lower free energy than any other state Achieved through kinetic pathway of unstable intermediates (not all intermediates are sampled) Assisted by chaperones and protein disulfide isomerases so intermediates are not trapped in a local low energy state Protein folding
The Kinetic Theory of Protein Folding • Folding proceeds through a definite series of steps or a Pathway. A protein does not try out all possible rotations of conformational angles, but only enough to find the pathway • - The final state may NOT be the most stable conformation possible, but it could be the most stable conformation that is accessible in a reasonable amount of time. This is also the biologically important time frame
Protein folding is initiated by reversible and rapid formation of local secondary structures Secondary structures then form domains through the cooperative aggregation of folding nuclei Domains finally form the final protein through “Molten globule” intermediates. Folding Pathways
Models for Protein Folding 1. Hydrophobic collapse. Formation of a 'molten globule' 2. Framework model. Secondary structure forms first, perhaps including supersecondary structure. 3. Nucleation. Domains fold independently, and sub-domains serve as 'structural kernels’.
Local interactions are the main determinants of protein structures Interactions as the ones responsible for forming secondary structural elements, a-helices and b-sheets Isolated helices/sheets form early in the protein folding pathway, then assemble in the native tertiary structure Framework model
Hydrophobic side chains are being buried Secondary structure formation insulates the polar protein backbone from the nonpolar protein interior Hydrogen bonds, disulfide bonds and salt bridges begin to form and stabilize structure van der Waals interactions bring protein substructures into stable contact What's really happening?
Nonspecific interactions (hydrophobic effect, van der Waals) are required to bring the protein together into a globular conformation Steric interactions (restraints on the backbone) limit the ways in which the globular conformation can form Chemically specific interactions (hydrogen bonds, ionic interactions, dipolar interactions) determine the fine detail of the protein structure What's most important in folding?
Enzymes that speed folding • Protein disulphide isomerase Facilitates formation of correct disulphide bridges • Peptidyl proline isomerase Catalyses cis-trans isomerisation of peptide bonds involving proline • Molecular chaperones Help folding, especially of large proteins, by preventing interaction with other proteins
What is the molecular basis of the observation that proteins of thermophiles are more stable than their homologues in mesophiles?