Download
1 / 41
410 likes | 536 Views
Generating the histone code CDK8 kinases histones to repress transcription. Generating the histone code CDK8 kinases histones to repress transcription Appears to interact with mediator to block transcription. Generating the histone code

E N D
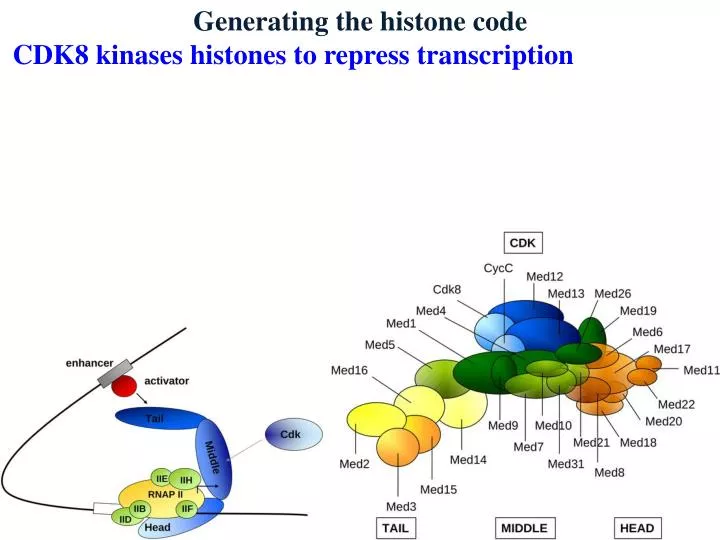
Generating the histone code • CDK8 kinases histones to repress transcription
Generating the histone code • CDK8 kinases histones to repress transcription • Appears to interact with mediator to block transcription
Generating the histone code • Rad6 proteins ubiquitinate histone H2B to repress transcription • Polycomb proteins ubiquitinate histone H2A to silence genes
Generating the histone code • CDK8 kinases histones to repress transcription • Appears to interact with mediator to block transcription • Phosphorylation of Histone H3 correlates with activation of heat shock genes!
Generating the histone code • CDK8 kinases histones to repress transcription • Appears to interact with mediator to block transcription • Phosphorylation of Histone H3 correlates with activation of heat shock genes! • Phosphatases reset the genes
Generating the histone code • Rad6 proteins ubiquitinate histone H2B to repress transcription
Generating the histone code • Rad6 proteins ubiquitinate histone H2B to repress transcription • Polycomb proteins ubiquitinate histone H2A to silence genes • A TFTC/STAGA module mediates histone H2A and H2B deubiquitination, coactivates nuclear receptors, and counteracts heterochromatin silencing
Generating the histone code • Many proteins methylate histones: highly regulated!
Generating the histone code • Many proteins methylate histones: highly regulated! • Methylation status determines gene activity
Generating the histone code • Many proteins methylate histones: highly regulated! • Methylation status determines gene activity • Mutants (eg Curly leaf) are unhappy!
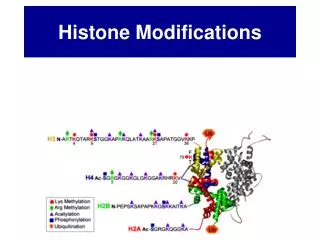
Generating the histone code • Many proteins methylate histones: highly regulated! • Methylation status determines gene activity • Mutants (eg Curly leaf) are unhappy! • Chromodomain protein HP1 can tell the difference between H3K9me2 (yellow) • & H3K9me3 (red)
Generating the histone code • Chromodomain protein HP1 can tell the difference between H3K9me2 (yellow) & H3K9me3 (red) • Histone demethylases have been recently discovered
Next step: deciding which genes to clone • Problem = correlating enzymes with genes • Who matches the pH? • Who localizes where? • Which isoform if alternatively spliced? • Clone several, using one known to work to find orthologs • Use sequence to design primers to clone cDNA
How to proceed? Kinetic and Spectroscopic Studies of Bicupin Oxalate Oxidase and Putative Active Site Mutants
Primer/probe design • Crucial for successful DNA & RNA analysis! • Main source of specificity for PCR • good primers only bind your sequence
Primer/probe design • Also important for microarrays, sequencing, Southerns • Concerns • Specificity: only want them to bind at one place • Main concern: 3’ end should not bind
Primer/probe design • Also important for microarrays, sequencing, Southerns • Concerns • Specificity • Complementarity: • Hairpins • homoduplexes • heteroduplexes • may not melt • May be extended by DNA polymerase
Primer/probe design • Also important for microarrays, sequencing, Southerns • Concerns • Specificity • Complementarity: • Melting T • Should match! • Every site calculates them differently!
Primer/probe design • Also important for microarrays, sequencing, Southerns • Concerns • Specificity • Complementarity: • Melting T • Targeting specific locations • amplifying specific sequences • creating mutations: need mismatch towards 5’ end so 3’ end binds well • Add restriction sites at 5’ end: may need to reamplify an amplicon
Choosing enzyme • Template • Fidelity • Taq from Thermus aquaticus has no proof-reading • goes faster, but error freq of 1 in 3000 • Vent from Thermococcus litoralis has error frequency of 1 in 30,000 • Pfu from Pyrococcus furiosus has error frequency of 1 in 400,000 • Genetically engineered proof-reading Phusion from NEB has error frequency of 1 in 2,000,000
Choosing enzyme • Template • Fidelity • Temperature stability • E.coli DNA polymerase I denatures at 75˚ C • T1/2 of Taq @ 95˚ C is 0.9 hours, < 0.1 hour @ 100˚ C • T1/2 of Phusion @ 96˚ C is >6 hours, 2 hours @ 98˚ C • T1/2 of Vent @ 95˚ C is 6.7 hours, 1.8 hours @ 100˚ C • T1/2 of Deep Vent from Pyrococcus species GB-D (grows @ 104˚ C)is 23 hours @ 95˚ C, 8 hours @ 100˚ C
Choosing enzyme • Template • Fidelity • Temperature stability • Processivity (how far does it go before falling off) • Phusion is 10x more processive than Pfu, 2x more than Taq • lets you make longer amplicons in shorter time • Taq = 8 kb max cf 40 kb for Phusion
Choosing enzyme • Template • Fidelity • Temperature stability • Processivity (how far does it go before falling off) • Km • dNTP: 13 µM for Taq, 60 µM for Vent • DNA: 2 nM for Taq, 0.01 nM for Deep Vent • Vmax: >1,000 nt/s when attached • Binding is limiting, processivity determines actual rate • 1000 bp/min is good for PCR • Tolerance of imperfect conditions • Dirty DNA: in general, non-proofreading polymerases tolerate dirtier DNA than proof-readers except Phusion
Choosing enzyme • Template • Fidelity • Temperature stability • Processivity (how far does it go before falling off) • Km • Vmax • Tolerance of imperfect conditions • Dirty DNA: in general, non-proofreading polymerases tolerate dirtier DNA than proof-readers except Phusion • dNTP analogs or modified dNTP • non-proofreading polymerases do better, but varies according to the modification • [Mg]: Vent is more sensitive to [Mg] and needs 2x more than Taq
Choosing enzyme • Template • Fidelity • Temperature stability • Processivity (how far does it go before falling off) • Km • Vmax • Tolerance of imperfect conditions • Fragment ends: proof-readers (eg Vent) give blunt ends • Non-proof-readers (eg Taq) give a mix of blunt & 3’A • Can use 3’A for t:A cloning • GAATTCAtcgca • CTTAAGtagcgt
Choosing enzyme • Template • Fidelity • Temperature stability • Processivity (how far does it go before falling off) • Km • Vmax • Tolerance of imperfect conditions • Fragment ends: proof-readers (eg Vent) give blunt ends • Cost @ NEB: http://www.neb.com/nebecomm/default.asp • Taq = $59.00 for 400 units • Vent = $62.00 for 200 units • Deep Vent = $90.00 for 200 units • Phusion = $ 103.00 for 100 units
Optimizing PCR • [enzyme] • [Template] • [Mg2+] • Annealing Temperature • Denaturation temperature
Optimizing PCR • [enzyme] • 0.4-2 units/100 µl for proofreaders : start with 1
Optimizing PCR • [enzyme] • 0.4-2 units/100 µl for proofreaders : start with 1 • 1-5 units/100 µl for non-proofreaders : start with 3
Optimizing PCR • [enzyme] • [Template] • 1-10 ng/100 µl reaction for plasmids • 10 - 1000 ng/100 µl reaction for genomic DNA • Excess DNA can give extra bands, also brings more contaminants
Optimizing PCR • [enzyme] • [Template] • 1-10 ng/100 µl reaction for plasmids • 10 - 1000 ng/100 µl reaction for genomic DNA • Excess DNA can give extra bands, also brings more contaminants • [dNTP] • 50-500 µM for Taq: start with 200, lower increases fidelity, higher increases yield • 200-400 µM for proof-readers: if too low start eating
Optimizing PCR • [enzyme] • [Template] • [Mg2+] • 0.5 - 4 mM for Taq: start with 1.5; lower if extra bands, raise if low yield
Optimizing PCR • [enzyme] • [Template] • [Mg2+] • 0.5 - 4 mM for Taq: start with 1.5; lower if extra bands, raise if low yield • 1- 8 mM for proofreaders: start with 2, lower if extra bands, raise if low yield
Optimizing PCR • [enzyme] • [Template] • [Mg2+] • Denaturation Temperature • Go as high as you can w/o killing enzyme before end • 94˚C for Taq • 96-98˚C for Vent • 98˚C for Deep Vent & Phusion
Optimizing PCR • [enzyme] • [Template] • [Mg2+] • Denaturation Temperature • Annealing Temperature • Start5 ˚C below lowest primer Tm
Optimizing PCR • [enzyme] • [Template] • [Mg2+] • Denaturation Temperature • Annealing Temperature • Start5 ˚C below lowest primer Tm • Adjust up and down as needed • Use temperature gradient feature to find best T
Optimizing PCR • [enzyme] • [Template] • [Mg2+] • Denaturation Temperature • Annealing Temperature • Start5 ˚C below lowest primer Tm • Adjust up and down as needed • # cycles: raise if no bands, lower if OK yield but extra bands
Optimizing PCR • Most common problems = wrong [DNA], dirty DNA, [Mg2+] annealing temperature & # cycles
Optimizing PCR • Most common problems = wrong [DNA], dirty DNA, [Mg2+] annealing temperature & # cycles • Can try “PCR enhancers” to overcome dirty DNA • Use Ammonium SO4 in buffer cf KCl • Use molecules that alter Tm eg DMSO & formamide • Use molecules that stabilise Taq eg Betaine & BSA
Optimizing PCR • Most common problems = wrong [DNA], dirty DNA, [Mg2+] annealing temperature & # cycles • If extra bands persist, use Taq bound to antibody • Inactive until denature antibody 7’ at 94˚ C
Optimizing PCR • Most common problems = wrong [DNA], dirty DNA, [Mg2+] annealing temperature & # cycles • If extra bands persist, use Taq bound to antibody • Inactive until denature antibody 7’ at 94˚ C • Alternatively, try touch-down: start annealing @ too high & lower 1˚ C each cycle ( binds correct target first)