Download

1 / 1
10 likes | 170 Views
CONCLUSIONS. Identification of the immunoreactive proteins in the SAM. Preparation of the C. rectus GroEL-like protein Purification of the C. rectus GroEL-like protein was performed according to the method of Hinode et al (7). C. rectus ATCC 33238 cells Heat shock at 44 o C for 4 h

E N D
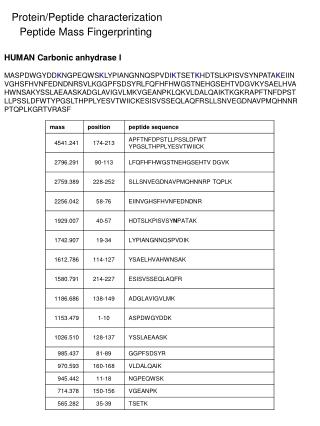
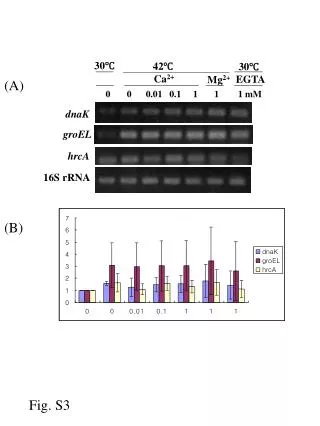
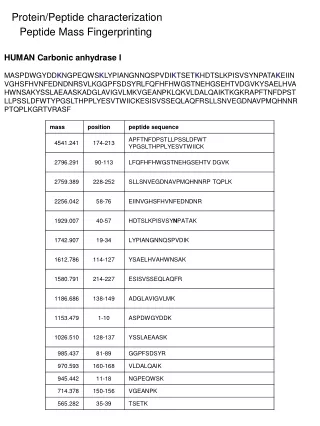
CONCLUSIONS Identification of the immunoreactive proteins in the SAM Preparation of the C. rectus GroEL-like protein Purification of the C. rectus GroEL-like protein was performed according to the method of Hinode et al (7). C. rectus ATCC 33238 cells Heat shock at 44oC for 4 h Suspend in cell lysis buffer and sonicate Centrifuge at 10,000 g for 20 min Collect supernatant and apply onto an adenosine 5’-triphosphate (ATP)-agarose column Elute with 5 mM ATP in the above buffer Collect fractions containing the GroEL-like proteins Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and electroelution Dialyze the eluted sample with distilled water and store at -20oC Preparation of specific HSP antibodies The purified C. rectus GroEL-like protein (10 µg) was injected intracutaneously into a New Zealand white rabbit in the presence of complete Freund’s adjuvant. Subsequent injections in the presence of incomplete Freund’s adjuvant were given intramuscularly at days 9, 20 and 41. The rabbit was bled via the heart directly at day 48. The antiserum (pAb-CrGroEL) was pooled and stored at -80oC until use. INTRODUCTION RESULTS METHODS Immunoreactivity of the C. rectus GroEL-like protein Fig. 1. Homogeneity of purified GroEL-like proteins After SDS-PAGE, 12% polyacrylamide gels were stained with silver nitrate. The amount of protein applied was 0.2 µg in each lane. Lane 1: Purified GroEL-like protein from C. rectus ATCC 33238 Lane 2: Purified GroEL-like protein from A. actinomycetemcomitans ATCC 29522 REFERENCES A homogenous protein with an apparent molecular mass of 64-kDa was detected in the purified preparations of GroEL from C. rectus ATCC 33238 as well as from A. actinomycetemcomitans. The purification procedure yielded 316.8 µg of C. rectus 64-kDa protein from 3-l culture. Using this purified protein for immunization, a high-titer serum of pAb-CrGroEL was obtained from each immunized rabbit. Fig. 4. Two-dimensional SDS-PAGE of the C. rectus SAM stained with silver nitrate (panel A) and Western immunoblotting probed with pAb-CrGroEL (panel B) Sixty micrograms of the SAM and the pI marker were added to a strip-gel in a pH range of 3-10 and separated initially by isoelectric focusing, then electrophoresed using 12% polyacrylamide gel. The four pI markers are shown with circles in panel A. Fig. 2. SDS-PAGE/Western immunoblotting of the C. rectus cells Panel A: Probed with pAb-CrGroEL Panel B: Probed with mAb-HuHSP60 Panel C: Probed with pAb-AaGroEL Panel D: Probed with preimmune rabbit serum Lanes 1, 3 and 5: Unstressed cells (37oC) Lanes 2, 4 and 6: Stressed cells (44oC) Lanes 1 and 2: C. rectus ATCC 33238 Lanes 3 and 4: C. rectus 325 Lanes 5 and 6: C. rectus 796 A 12% polyacrylamide gel was used and 10 µg of protein of the solubilized cells was applied in each lane. Two proteins from the prepared SAM were visualized by two-dimensional Western immunoblotting with pAb-CrGroEL (Fig. 4, panel B): the 150-kDa protein with a pI of approximately 4.8 (large arrow) and the 64-kDa protein with a pI of approximately 4.7 (small arrow). In order to identify the 150-kDa protein, 20 cycles of Edman degradation were carried out on the sample. The N-terminal amino acid sequence was found to be AALTQ TQVGQ LYVRL FGRVD. This sequence revealed a strong homology (80%) with the deduced amino acid sequence of the crs gene (crs: the gene encoding C. rectus S-layer protein), MALTQ TQVSQ LYVTL FGRVS (8). When the alignment of the deduced amino acid sequence of C. rectus GroEL-like protein was compared with that of crs homolog, little homology was revealed (data not shown). The purified GroEL-like protein from Actinobacillus actinomycetemcomitans ATCC 29522 and its specific polyclonal antibody (pAb-AaGroEL) were also used and prepared as above. A monoclonal antibody directed to the recombinant human HSP60 (mAb-HuHSP60) was purchased from StressGen. Electrophoresis and Western immunoblotting SDS-PAGE Transfer onto a nitrocellulose membrane Block with 3% bovine serum albumin (BSA) Incubate with specific HSP-antibodies as a first antibody Incubate with second antibodies coupled to horseradish peroxidase (HRP) Develop with HRP color development reagent Observation by transmission electron microscopy (TEM) Wash C. rectus cells with PBS Fix with 10% formaldehyde solution Place in PBS containing 1% BSA Incubate with HSP-antibodies Immunogold labeling Apply to coated mesh copper grids and stain with 2% uranyl acetate Observe by TEM Biochemical nature of antigenic determinants The biochemical nature of antigenic determinants recognized by pAb-CrGroEL was investigated using two proteolytic enzymes and periodic acid. Incubate the SAM or the GroEL-like protein with 0.5% protease (from Streptomyces griseus) or 0.1% trypsin (from bovine pancreas) at 37oC for 30 min Stop the reaction by boiling for 10 min SDS-PAGE/Western immunoblotting Incubate the SAM or the GroEL-like protein with 1% periodic acid at room temperature for 30 min Neutralize by the addition of NaOH solution SDS-PAGE/Western immunoblotting Figure 2 shows the reactive bands of C. rectus cells with specific antibodies. No bands (panel D) were observed when 1:3,000 dilution was used. All strains of C. rectus possessed immunoreactive bands at 64-kDa not only against pAb-CrGroEL (panel A) but also mAb-HuHSP60 (panel B) and pAb-AaGroEL (panel C). In some cases, increased synthesis of the GroEL-like protein under heat-stress was found. Besides the 64-kDa protein, strain ATCC 33238 strain showed an immunoreactive band at 150-kDa against pAb-CrGroEL (panel A, lanes 1 and 2). The 150-kDa band in C. rectus ATCC 33238 was not detected when mAb-HuHSP60 (panel B, lanes 1 and 2) or pAb-AaGroEL (panel C, lanes 1 and 2) was used as the first antibody. Biochemical nature of the immunoreactive epitopes TEM observation Fig. 3. Immunogold labeling with specific HSP-antibodies in C. rectus cells Panel A: ATCC 33238 with pAb-CrGroEL (1:200 dilution) Panel B: 325 with pAb-CrGroEL (1:200 dilution) Panel C: 796 with pAb-CrGroEL (1:100 dilution) Panel D: ATCC 33238 with mAb-HuHSP60 (1:4 dilution) Panel E: ATCC 33238 with pAb-AaGroEL (1:100 dilution) Panel F: ATCC 33238 with preimmune rabbit serum (1:100 dilution) Bar = 0.5 µm Fig. 5. Characterization of the immunoreactive epitopes found on the C. rectus SAM and on the C. rectus GroEL-like protein Panel A: Immunoblotting of the C. rectus SAM (5 µg/lane) probed with pAb-CrGroEL Panel B: Immunoblotting of the C. rectus GroEL-like protein (0.07 µg/lane) probed with pAb-CrGroEL Panel C: Immunoblotting of the C. rectus GroEL-like protein (0.07 µg/lane) probed with mAb-HuHSP60 In immunogold labeling with specific pAb-CrGroEL, many gold beads were detected on the cell surface of C. rectus ATCC 33238 (Fig. 3, panel A), while few were found on the cells of strain 325 (panel B) and strain 796 (panel C). No difference between pAb-AaGroEL gold beads (panel E) and preimmune gold beads (panel F) was observed. No mAb-HuHSP60 gold beads were detected on the cells of ATCC 33238 (panel D) as well as the other strains (data not shown). Lane 1: No treatment Lane 2: Treatment with 0.5% protease (from Streptomyces griseus) Lane 3: Treatment with 0.1% trypsin (from bovine pancreas) Lane 4: Treatment with 1% periodic acid All panels were transferred from 7.5% polyacrylamide gels. The same amount of protein was used in all cases before and after treatment. The migration front is indicated by the open arrow. 0731 To characterize the biochemical nature of immunoreactive epitopes found in the SAM and on the GroEL-like protein, SDS-PAGE/Western immunoblotting were performed after treatment with either periodic acid or proteolytic enzymes. The 64-kDa GroEL-like protein was completely degraded when the sample was treated with proteolytic enzymes (Fig. 5, panels B and C, lanes 2 and 3). However, the samples treated with 1% periodic acid reacted strongly with pAb-CrGroEL as well as with mAb-HuHSP60 (panels B and C, lane 4). These facts suggested that the immunodominant epitopes of the C. rectus GroEL-like protein were peptidic in nature. When the 150-kDa protein in the SAM was treated with proteolytic enzymes, several digested bands were detected (panel A, lanes 2 and 3), whereas a treatment with periodic acid led to only a faint band detected at the migration front (panel A, lane 4). Recently, considerable attention has been focused on the potential role of bacterial heat shock proteins (HSPs) in the disease process of bacterial infections. HSPs synthesized by both prokaryotic and eukaryotic cells are highly conserved, and are classified into families according to their molecular mass. It is thought that HSPs are expressed under stressed conditions during an infection process when host and pathogen interact. We previously reported that specific antibodies directed against HSPs from periodontopathogenic bacteria cross-reacted with those of human origin (1). It was also found that gingival homogenate samples from patients with adult periodontitis reacted with antihuman HSP60 (2). Furthermore, several studies have shown a positive relationship between the severity of periodontal disease and the serum antibody titer to HSPs (3,4,5). These observations support the hypothesis that HSPs could trigger an immune response involved in tissue damage associated with the disease. Campylobacter rectus is a gram-negative, asaccharolytic and agar-corroding bacteria isolated from lesions with advanced destructive periodontitis. C. rectus possesses a surface layer (S-layer) protein, which is considered a putative virulence factor. Kokeguchi et al. (6) reported that IgG titers directed to the S-layer protein in sera from patients with periodontitis were higher than those in healthy donors. In this study, we investigated the antigenic characteristics of the GroEL-like protein (HSP60 family) from C. rectus. Immunological Characterization of the GroEL-Like Proteinfrom Campylobacter rectus M. YOKOYAMA*, S. TANABE, M. YOSHIOKA, D. HINODE and R. NAKAMURADepartment of Preventive Dentistry, School of Dentistry, The University of Tokushima, Japan The GroEL-like protein from C. rectuswas characterized immunologically and the following results were obtained: 1) A homogenous protein with an apparent molecular mass of 64-kDa was detected in the purified preparations of GroEL from C. rectus ATCC 33238 as well as from A. actinomycetemcomitans. 2) PAb-CrGroEL was reactive also with the 150-kDa protein. 3) This 150-kDa protein was found to be present on the surface-associated material of C. rectus. 4) Analysis of the first 20 N-terminal amino acid sequence of the 150-kDa protein revealed a strong homology (80%) with that of the C. rectus S-layer protein. 5) The immunoreactive epitopes were supposed to be carbohydrate. Results described above suggested that the GroEL-like protein and the S-layer protein of C. rectus may share the same carbohydrate epitopes as an antigenic determinant. C. rectus strains and growth conditions Campylobacter rectus ATCC 33238, 325 (clinical isolate) and 796 (clinical isolate) weregrown in trypticase soy broth supplemented with 0.2% yeast extract, 0.3% phytone peptone, 0.2% NaCl, 0.3% ammonium formate, 0.4% sodium fumarate and 0.4% L-asparagine adjusted to pH 7.8. Bacterial cultures were incubated at 37oC in an anaerobic chamber (85% N2, 10% H2 and 5% CO2) and grown to mid-log phase (24-36 h incubation). Bacteria were transferred to sterile glass tubes in an anaerobic chamber and sealed with caps. Tubes were removed from anaerobic chamber and heat-shocked in a water bath at 37oC or 44oC for 6 h. After incubation, C. rectus cells were chilled in ice, washed with phosphate-buffered saline (PBS) and placed in 10% trichloroacetic acid at 4oC overnight. The bacterial cells were washed twice with PBS and stored at -20oC. Preparation of the surface-associated material (SAM) The unstressed cells of C. rectus ATCC 33238 were washed once with PBS, and then the SAM was extracted by gentle stirring in saline for 1h at 4oC. Bacterial cells were removed by centrifugation at 10,000 g for 20 min. The supernatant was dialyzed overnight against distilled water, lyophilized and stored at -20oC. 1. Hinode D et al. Oral Microbiol Immunol 1998: 13: 55-58. 2. Ando T et al. Microbiol Immunol 1995: 39: 321-327. 3. Lopatin DE et al. J Periodontol 1999: 70: 1185-1193. 4. Schett G et al. Int Arch Allergy Immunol 1997: 114: 246-250. 5. Tabeta K et al. Clin Exp Immunol 2000: 120: 285-293. 6. Kokeguchi S et al. J Clin Microbiol 1989: 27: 1210-1217. 7. Hinode D et al. J Microbiol Methods 1996: 25: 349-355. 8. Wang B et al. Infect Immun 1998: 66: 1521-1526.