Download

1 / 44
480 likes | 716 Views
DNA Replication. “A structure this pretty just had to exist.” —James Watson, The Double Helix,1968. PART I The complete story. How DNA was found to be the hereditary material??. DNA: The Genetic Material. Mendel’s experiments showed that genes were the basic unit of inheritance.

E N D
DNA Replication “A structure this pretty just had to exist.” —James Watson, The Double Helix,1968
PART IThe complete story • How DNA was found to be the hereditary material??
DNA: The Genetic Material Mendel’s experiments showed that genes were the basic unit of inheritance. Genes are located on chromosomes Chromosomes= DNA + Proteins So question arises?? Which Chemical component is the Genetic material? Protein OR DNA??
Fredrick Griffith Experiment 1920s Transforming Principle
The S strain DNA contains the genes that form the protective polysaccharide capsule. While the bacteria had been killed, the DNA had survived the heating process and was taken up by the R strain bacteria. • Griffiths Conclusion: Some material in the heat-killed S strain that was responsible for transforming the R-strain into a lethal form Transformation = passing the inheritance factor from one organism to another THIS TRANSFORMING PRINCIPLE WAS LATER FOUND TO BE DNA
Oswald Avery: He had reported that DNA, not protein (which was believed at the time), was the hereditary substance (transforming material). • he treated samples known to contain the pneumococcal transforming principle to destroy different types of molecules—proteins, nucleic acids, carbohydrates, and lipids—and tested the treated samples to see if they had retained transforming activity
. The answer was always the same: If the DNA in the sample was destroyed, transforming activity was lost, but there was no loss of activity when proteins, carbohydrates, or lipids were destroyed
TO COFIRM WHETHER DNA OR PROTEIN IS THE HEREDITARY MATERIAL • The Hershey–Chase Experiment • Using T2 bacteriophage :virus that infects bacteria, consists of little more than a DNA core packed inside a protein coat • When a T2 bacteriophage attacks a bacterium, part of the virus enters the bacterial cell. About 20 minutes later, the cell bursts, releasing dozens of viruses
The entry of a viral component changes the genetic program of the host bacterial cell: it is converted from a bacterium into a bacteriophage factory. Hershey and Chase set out to determine which part of the virus protein or DNA—entersthe bacterial cell
The Hershey–Chase Experiment S35 LABELLED P32 LABELLED
One batch of T2 viruses grown in presence of S35 had S35 in their protein coat. The other batch of T2 viruses grown in presence of P32 had P32 in their DNA.
PART II DNA STRUCTURE • How DNA structure was established??
Chargaff’s rule • The amount of adenine equals the amount of thymine (A =T), and the amount of guanine equals the amount of cytosine (G = C). As a result, the total abundance of purines (A + G) equals the total abundance of pyrimidines (T + C).
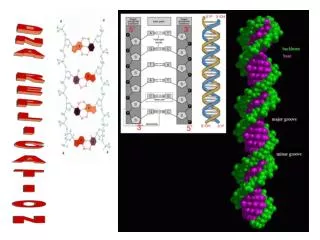
Watson and Crick Model of DNA Structure Key features of DNA Double stranded Uniform diameter Right handed helix twists like a Screw Anti parallel Hydrogen bonding Complementary base pairing
DNA Structure- a revision • Nucleotide • Nucleoside • Purines • Pyrimidines
Anti parallel strands The 5'-end (pronounced "five prime end") designates the end of the DNA or RNA strand that has the fifth carbon in the sugar-ring of the deoxyribose or ribose at its terminus (the end with the free phosphate) The 3'-end of a strand is so named due to it terminating at the hydroxyl group of the third carbon in the sugar-ring, and is known as the tail end(the end with the free hydroxyl)
-diester refers to the two bonds formed by -OH groups reacting with phosphate groups the phosphodiester bond ester phosphodiester
structure • In both RNA and DNA, the backbone of the macromolecule consists of alternating pentose sugars and phosphates (sugar—phosphate—sugar—phosphate). The bases are attached to the sugars and project from the chain.
The nucleotides are joined by phosphodiester linkages between the sugar of one nucleotide and the phosphate of the next .The phosphate groups link carbon 3 in one pentose sugar to carbon 5 in the adjacent sugar.
Concept of complementarity • The two anti parallel polynucleotide chains of double-helical DNA are not identical in either base sequence or composition. Instead they are complementary to each other • Every base pair consists of one purine (A or G) and one pyrimidine (T or C). This pattern is known as complementary base pairing. Because the AT and GC pairs are of equal length and fit identically into the double helix, the diameter of the helix is uniform • information contained in the sequence of one strand is conserved in the sequence of the other
Why does A bond only with T and C only with G? What do you notice? • Nitrogenous Bases T and C are single-ring Pyrimidines . • A and G are double-ring Purines • A single ring base bonds with a double
PART III DNA REPLICATION • How identical copies of DNA are produced??
Determining the mechanism of DNA replication Three models: • Conservative • Semi Conservative • Dispersive
Can be explained only by the semi conservative model of DNA replication. • In the first round of DNA replication, the strands of the double helix—both heavy with 15N ,separated. Each strand then acted as the template for a second strand, which contained only 14N and hence was less dense. Each double helix then consisted of one 15 N strand and one 14 N strand, and was of intermediate density • In the second replication, the 14 N-containing strands directed the synthesis of partners with 14 N, creating low-density DNA, and the15 N strands formed new 14 N partners
In conservative replication, the first generation would have had both high-density DNA (15 N–15N) and low-density DNA (14N–14N), but no intermediate-density DNA. • In dispersive replication, the density of the new DNA would have been half that of parental DNA
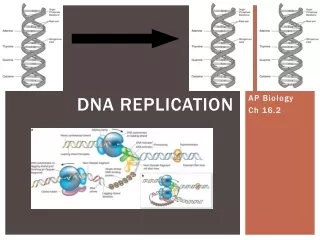
DNA replication Replication:process of producing two identical replicas from one original DNA molecule. Each strand of the original DNA molecule serves as template for the production of the complementary strand • a huge protein complex called the replication complex • the replication complex seems to be stationary, it is the DNA that moves
DNA replicates in both directions from the origin of replication (ori), forming two replication forks • Small circular chromosomes, such as the 3-million-base pair DNA of bacteria, have a single origin of replication. Human chromosome with 80 million base pairs, there are hundreds of origins of replication.
DNA Replication Mechanism • Initiation • DNA helicase uses energy from ATP hydrolysis to unwind the DNA • special proteins called single strand binding proteins bind to the unwound strands to keep them from reassociating into a double helix • DNA polymerases cannot start a strand from scratch. Therefore, a short single strand of RNA, called a primer, is required for replication. This RNA strand, is synthesized by an enzyme called primase (RNA Polymerase)
Elongation: • DNA polymerase then adds nucleotides to the 3′end of the primer and continues until the replication of that section of DNA has been completed. • Then the RNA primer is degraded, DNA is added in its place, and the resulting DNA fragments are connected by the action of ligases
nucleotides are always added to the growing strand at the 3′end— the end at which the DNA strand has a free hydroxyl (—OH) group on the 3′ carbon of its terminal deoxyribose The three phosphate groups in a deoxyribonucleoside triphosphate areattached to the 5′position of the sugar. So when a new nucleotide is added to DNA, it can attach only to the 3′end
When DNA polymerase brings a deoxyribonucleoside triphosphate with the appropriate base to the 3′ end of a growing chain, the free hydroxyl group on the chain reacts with one of the substrate’s phosphate groups.. The phosphate group still on the nucleotide becomes part of the sugar–phosphate backbone of the growing DNA molecule.
Termination: • Eukaryotes initiate DNA replication at multiple points in the chromosome, so replication forks meet and terminate at many points in the chromosome. • Termination requires that the progress of the DNA replication fork must stop or be blocked. Involves the interaction between two components: (1) a termination site sequence in the DNA, and (2) a protein which binds to this sequence to physically stop DNA replication,named the DNA replication terminus site-binding protein, or Ter protein
single strand binding proteins Helicase
summaryENZYMES FOR DNA REPLICATION • Helicase = separates 2 DNA strands (breaks H bonds) • Primase =RNA primers at INITIATION • Topoisomerase = unwinding DNA • DNA Polymerase = Adding of DNA nucleotides • LIGASE = Binds the Okazaki fragments • single strand binding proteins =bind to the unwound strands to keep them from reassociating into a double helix
The DNA polymerases are only able to “read” the parental nucleotide sequences in the 3'→5' direction, and they synthesize the new DNA strands only in the 5'→3' (antiparallel) direction. Therefore, the two newly synthesized stretches of nucleotide chains must grow in opposite directions—one toward the replication fork and one away from the replication fork .
1. Leading strand: The strand that is being copied in the direction of the advancing replication fork is called the leading strand and is synthesized continuously. • 2. Lagging strand :The strand that is being copied in the direction away from the replication fork is synthesized discontinuously, with small fragments of DNA being copied near the replication fork. These short stretches of discontinuous DNA, termed Okazaki fragments, are eventually joined (ligated) to become a single, continuous strand.
telomeres • Telomeres are complexes of noncoding DNA plus proteins located at the ends of linear chromosomes. They maintain the structural integrity of the chromosome, preventing attack by nucleases
Following removal of the RNA primer from the extreme 5'-end of the lagging strand, there is no way to fill in the remaining gap with DNA. Consequently, in most normal human somatic cells, telomeres shorten with each successive cell division. Once telomeres are shortened beyond some critical length, the cell is no longer able to divide and is said to be senescent. In germ cells and other stem cells, as well as in cancer cells, telomeres do not shorten and the cells do not senesce. This is a result of the presence of telomerase, which maintains telomeric length.