Download

1 / 127
1.29k likes | 1.67k Views
Mechanisms of Microevolution. Reading:Freeman, Chapter 24, 25. Microevolution vs. Macroevolution. The term “ microevolution ” applies to evolutionary change within a lineage It occurs continuously.

E N D
Mechanisms of Microevolution Reading:Freeman, Chapter 24, 25
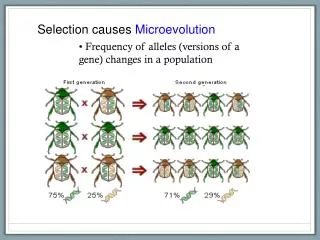
Microevolution vs. Macroevolution • The term “microevolution” applies to evolutionary change within a lineage • It occurs continuously. • Depending upon the organism and the circumstances, it can transform a lineage. dramatically over time. • Alternately, a lineage may appear to remain the same over time-this is called stasis. • Macroevolution is the origin and extinction of lineages. • It can happen gradually, or slowly. • Both processes are essential to evolution. Microevolution is probably better understood, and better documented, because in some organisms it takes place on timescales we can study directly by experiment and direct observation. • Ironically, in “On the Origin of Species” Darwin lays out a theory of microevolution…he assumed macroevolution would inevitably result from microevolution. • It would be 100 years later that Ernst Mayr, and others, would develop a scientific theory of speciation. • The replacement of one species by another (as opposed to the replacement of one allele by another), by the way, is an ecological process..it is not evolution in the usual sense, though this phenomenon usually leads to extinction of some species and diversification of others.
The Population is the Basic Unit of Microevolutionary Change • The genotype of an individual is, essentially, fixed at birth. • The population is the smallest unit where evolutionary change is possible. • Unlike individuals, populations permit the origin of new alleles through mutation, and the change in the frequency of alleles through selection, genetic drift, etc.. • Individuals do not evolve, populations and species evolve.
Population Genetics • Population genetics refers to the study of evolution via the observation and modeling of allele frequencies and genetic change in populations of organisms. • There are three parameters to keep in mind: • allele frequency: the proportion of a specific allele at a given locus, considering that the population may contain from one to many alleles at that locus. • genotype frequency: the proportion of a specific genotype at a given locus, considering that many different genotypes may be possible. • phenotype frequency: the proportion of individuals in a population that exhibit a given phenotype.
Consider a population of N organisms. • Two phenotpyes, yellow and tan. • Suppose that they are diploid and reproduce sexually. • Consider one gene with two alleles, A and a. • The possible genotypes are therefore: • AA, Aa, and aa.
Phenotype Frequencies • To calculate the frequency of a phenotype, count the number of individuals with that phenotype, and divide by the total. Therefore, the frequency of the yellow phenotype in the population below is 4/10=.40
Genotype Frequencies- • To calculate the frequency of a genotype in the population, find the total number of individuals in the population with that genotype, and divide by the population size, N. • f(AA)= #(AA)/N • f(Aa)= #(Aa)/N • f(aa)= #(aa)/N
Question: What are the frequencies of the AA and Aa, and aa genotypes in the population below? Aa Aa aa aa AA Aa aa Aa aa AA
Answer: freq(AA)=2/10=.20 freq(Aa)=4/10=.40 freq(aa)=4/10=.40 Aa Aa aa aa AA Aa aa Aa aa AA
Allele Frequencies- • By convention, the frequency of the dominant allele is called p, thus the frequency of the recessive allele, q=1-p. • To calculate the frequency of an allele in the population, add the total number of homozygotes for that allele to half the heterozygotes, and divide by the population size, N. • p= ((#AA) + (1/2)(#Aa))/N • q= ((#aa) + (1/2)(#Aa))/N • If you already know the genotype frequencies, • p=f(AA)+(1/2)f(Aa) • q=f(aa)+(1/2)f(Aa)
Question: What are the frequencies of the A and a alleles in the population below? Aa Aa aa aa AA Aa aa Aa aa AA
Answer: freq(A)=p=(4+(2x2))/20=.40 • freq(a)=q=(4)+(4x2)/20=.60 or q=1-.4 • since p+q=1 Aa Aa aa aa AA Aa aa Aa aa AA
Evolutionary Change is a Consequence of Changes in Allele Frequencies • This is the genetic definition of evolution…a synthesis between Mendel’s and Darwin’s theories. • All of the evolutionary change between our single-celled ancestors and ourselves can be described as the sequential origin of new alleles, their replacement of old ones, and occasionally the origin of new genes through duplication..
Evolution as Change in Allele/Genotype Frequency • Microevolutionary change is inherent in the change in the frequencies of alleles over time. • To be able to see if evolution IS occurring, we need to consider what we would expect if evolution is NOT occurring. • Once we know that, then if we see a departure, we know that evolution is occurring.
The Hardy-Weinberg Equilibrium • Hardy-Weinberg Equilibrium is defined as the situation in which no evolution is occurring. It is a genetic equilibrium. • It was the solution to a nineteenth century misconception-the notion that the dominance or recessiveness of an allele alone could cause evolutionary change (it can’t). • The Hardy-Weinberg Equilibrium refers to a particular locus: one locus may be undergoing rapid allele-frequency change, while other loci are in equilibrium.
Assumptions of the Hardy-Weinberg Equillibrium • A locus with two or more alleles will be in Hardy-Weinberg Equilibrium if five assumptions are met. These are: • 1. infinite population size (there are infinitely many individuals in the population.) • 2.there is no allele flow (i.e., no movement of individuals from population to population.) • 3.there is no mutation (no biochemical changes in DNA that produce new alleles.) • 4. there is random mating (this means that with regard to the trait we're looking at, individuals mate at random they don't select mates based on this trait in any way.) • 5. No Selection: the different genotypes (for the genetic trait we're studying) have equal fitness.
Consider a population of diploid, sexually reproducing individuals. Imagine a gene with two alleles, A and a, so there are three genotypes, AA, Aa, and aa. • Assume this population meets the five assumptions of Hardy-Weinberg Equilibrium. • Because mating is random, alleles mix together at random. • Because the population is infinitely large, the probability of getting a gamete with a particular allele in it is simply the frequency of that allele. • Similarly, determining the probabilities of getting particular genotypes will tell us thefrequencies of those genotypes in the population.
To get AA, you need an egg with allele A, with probability p, and a sperm with allele A, also with probability p. • The probability of getting both of these is p2 • Similarly, for aa, the chance of a sperm with an "a" allele is q and the chance of an egg with an "a" allele is also q, so: q2
There are two ways of getting the heterozygous genotype, Aa. These are: • 1.an "A" bearing sperm and an "a" bearing egg • 2.an "A" bearing egg and an "a" bearing sperm • The probability of an "A" bearing sperm is p; the probability of an "a" bearing egg is q, so the probability of the first way of getting an Aa is (p)(q) = pq. • Similarly, the probability of aA (a sperm, A egg) is also (p)(q)=pq. • So to get the probability of getting the Aa genotype, we add together the probabilities of the two ways of getting this genotype. So: The genotype frequency of Aa is pq+pq=2pq
With a little more math, it can be demonstrated that:As long as the conditions of the Hardy-Weinberg equilibrium are met, allele frequencies will remain constant. • After one round of random mating, there will exist a stable, mathematical set of genotype frequencies for any given allele frequencies. • This relationship will hold as long as the conditions of the Hardy-Weinberg equilibrium are met. • This means that if you know allele frequencies, you can solve for expected genotype frequencies. • If you know the frequency of a recessive genotype, you can usually infer the expected frequency of the recessive allele as well.
These apply under Hardy-Weinberg equilibrium: Freq(AA)=p2 Freq(Aa)=2pq Freq(aa)=q2 p2+2pq+q2=1
So-Without the HW equilibrium, you can do thus: • Allele frequencies from numbers of individuals • p= ((#AA) + (1/2)(#Aa))/N • q= ((#aa) + (1/2)(#Aa))/N • Genotype frequencies to allele frequencies • p=f(AA)+(1/2)f(Aa) • q=f(aa)+(1/2)f(Aa) • To go from allele frequency to genotype frequency, you must assume HW equilibrium • If this is not the case, there may be many potential genotype frequencies for a single allele frequency. • For instance, 500AA 0Aa 500aa, and 1000Aa both have p=.5, q=.5 • If HW is the case, use the following to go from genotype frequency to allele frequency, but remember these are estimates (and expected values of each would be these estimates times the total population size), not the true values: • Freq(AA)=p2 Freq(Aa)=2pq Freq(aa)=q2 • If you have the frequency of a recessive phenotype, its frequency is q2, thus, take the square root and get q. Get p from there, etc.
Question: • The albino color in rabbits is caused by a recessive allele. Aa and AA individuals are normally pigmented, and aa individuals are albino. • Imagine a population with 9999 normally pigmented rabbits, and a single albino rabbit. • What is the frequency of the recessive phenotype? What is the frequency of the (aa) genotype? • Under Hardy-Weinberg equilibrium, what are the expected frequencies of the dominant and recessive alleles? What is the expected frequency of the (Aa) genotype?
Answer. • Freq Recessive phenotype 1/10000=.0001 • Freq (aa) genotype, 1/10000, .0001 • q2=.0001, therefore q=(.0001)1/2 q=.01 • p+q=1, so p=1-.01=.99 • 2pq=(.01x.99x2)=.0198 • Note that while 1 in 10,000 rabbits is an albino, approximately one in fifty individuals carry the allele. This is a fairly common situation for a recessive allele which is selected against.
We can define five different ways in which evolution occurs based on the situations in which these five assumptions are NOT met. These are: • 1.genetic drift is change in allele frequency by random chance. It occurs if a population is not infinite in size. In populations that are not infinitely large, there will be random error in which alleles are passed from generation, and allele frequencies will change at random. Since no population is really infinitely large, there is always some genetic drift occurring; however, the effect is very small in large populations. The effect of genetic drift is larger in small populations.
2. Allele flow is change in allele frequency that occurs because individuals move among populations. If there are different allele frequencies in different populations of a species, then when individuals move to a new population, they will change theallele frequencies in the new population.
3. mutation is biochemical change in DNA that one allele into another and creates alleles. It not a common event (typical mutation rates are about one mutation in a million genes passed from generation to generation ); as a result, evolution through mutation is extremely slow. • Mutation is very important for evolution because, ultimately, mutation is the source of genetic variation. Other forms of evolution on cannot occur without genetic variation. • With regard to the fitness of alleles, mutation is random -- it may produce alleles that result in high fitness (rare) or low fitness (much more common), and the probability of a mutation is independent of an evolutionary “need” for the mutation.
4.non-random mating is evolution that occurs because individuals select mates based on their characteristics. • 5.natural selection is evolution that occurs because different genotypes have different fitness. More about this later.
You can test to see whether a population is in Hardy-Weinberg equilibrium, if you have the numbers of individuals of each genotype. -The Chi-Square test is ideal for this: -Generate expected values from HW expectations and compare them to observed values. -The “null hypothesis” in this case is that the population IS in HW equilibrium, and thus, no evolution is ocuring at that locus at the moment. -A violation might signal that some force of evolution is at work.
For instance, imagine that you had a population of river grapes Vitis riparia. • There is an enzyme locus you find interesting,(could be anything, superoxide dismutase for instance). • It has two codominant alleles, SODF, for fast (because DNA fragments of it migrate rapidly on a gel), and SODS for slow, because DNA fragments migrate less quickly on a gel. • You don’t actually know much about evolution in this species, and you want to know if this locus is evolving. • You census 1000 individuals (at least you think so, they are grapes). • You get: • 460 SODFSODF • 23 SODFSODS • 517 SODSSODS • Is the population in HW equilibrium?
So, there is no dominant locus, but we will call the frequency of SODF p. • p=(460+11.5)/1000=.47, thus q=.53 • Observed Expected • 460 SODFSODF 221 SODFSODF • 23 SODFSODS 498 SODFSODS • 517 SODSSODS 281 SODFSODS • Thus Chisquare=approximately 910 • There is only one degree of freedom here (crazy, I know it seems, but we generated three expected values from just p and q, so df=1) • This is much lager than the critical value, so the locus is not in HW equilibrium. • Why? • A statistical test cannot answer that…but there are not enough heterozygotes. This (as you will learn later) is the signature of inbreeding, and grape plants self-fertilize. • Alternately, since grapes reproduce asexually as well as sexually, this could be the differential success of two genotypes, both homozygotes, at reproducing asexually.
The Mechanisms of Evolution • Mutation • Allele (gene) flow • Selection • Genetic Drift • Nonrandom mating • each one is, in essence, the result of a violation of one or more of the assumptions of the Hardy-Weinberg equilibrium
Mutation • A mutation is a change in the organism’s DNA. • Mutations may affect somatic (nonreproductive tissue), or they may affect the germ line (reproductive tissue). Except in clonal organisms, somatic mutations cannot generally be passed on. • Evolutionary biologists are interested in heritable mutations, the kind that can be passed on to the next generation. • A heritable mutation changes one allele into another, sometimes creating an allele that is not already present in the population.
Some mutations create dominant alleles, some create recessive or codominant alleles. • Some mutations are harmful or lethal, many are totally neutral-they have no effect, a few are favorable. • Whether a mutation is harmful, neutral, or favorable, depends upon its environment
Some types of mutations. • Substitution: one nucleotide is substituted for another, frequently this causes no change in the resulting organism, sometimes the change can be dramatic. • Insertion: DNA is inserted into a gene, either one nucleotide or many. Sometimes, entire genes are inserted by viruses and transposable elements. • Deletion: DNA bases are removed. • Small insertions and deletions can inactivate large stretches of a gene, by causing a frame shift that renders a gene meaningless. • Duplication: an entire gene is duplicated. • Transposition: DNA is moved to a new place in the genome, frequently this happens because of errors in meiosis or transposable elements.
Mutations are random events: their occurrence is independent of their selective value - i.e., they do not occur when they are needed any more often than they would otherwise. • Mutations at any single locus are rare events: mutation rates at a typical locus are about 1 in 106 gametes.
Since each individual has thousands of alleles, the cumulative effect of mutations is considerable: • Consider that each of us has about 3.5x104 loci, and the mutation rates are about 1x10-6 per locus, thus, about 1 in 30 of our gametes has a new mutation somewhere in its genome. That means about 7% of us are mutants, more or less. YOU could be a mutant.
Mutations are the ultimate source of genetic variation • Mutations are the only source of new alleles (other than the occasional transfer of alleles by viruses). • Mutation is thus the ultimate source of genetic variation…it creates the raw material upon which natural selection acts.
Example-an interesting mutation: • In humans, one interesting mutation is called the CCR-d32 allele (the locus is named CCR, it is one of many alleles at that locus) • This allele codes for a 32 base pair deletion that makes the protein nonfunctional. • Lacking this protein on the surface of their blood cells, homozygous individuals (it is effectively codominant) are essentially resistant to HIV-HIV cannot infect their cells. • This mutation did not arise because of HIV, best we can figure, it predates the evolution of HIV by hundreds or thousands of years, and was neutral (or possibly maintained by selection induced by the bubonic plague) until HIV entered out species!
Genetic Drift • Genetic drift is the change in allele frequencies that occurs by chance events. In essence, it is identical to the statistical phenomenon of sampling error on an evolutionary scale. • It is a random (stochastic) process. • Because sampling error is greatest in small samples and smallest in large samples, the strength of genetic drift increases as populations get smaller.
Effects of Genetic Drift • Does not generally result in adaptation. • In a large population - it has little effect unless enormous spans of time are involved. • (if vast spans of time ARE involved, the cumulative effects of drift on any species can be considerable-genetic drift is the primary mechanism for the substitution of neutral alleles over time, which is the mechanism of the molecular clock used in systematics.)
In a small population • alleles can be lost (usually the rare ones) • other alleles are fixed-their frequency reaches 1.0 • genetic variation is lost, resulting in at population can become homozygous at many loci
Founder Effect • The founder effect is genetic drift that occurs when when a few individuals, representing a fraction of the original allele pool, invade a new area and establish a new population.
Examples: • California Cypress-a very large population was established from a small number of individuals. • Founder effect occurs in many introduced species. • Amish-a religious minority, which is essentially an isolated population, established from a relatively small number of individuals.
Bottlenecks • Bottlenecks are periods of very low population size or near extinction. This is another special case of genetic drift. • The result of a population bottleneck is that even if the population regains its original numbers, genetic variation is drastically reduced
Examples: • Cheetahs -nobody knows exactly why it occurred, but cheetahs underwent an extreme population bottleneck several thousand years ago. As a result, they have very little genetic variation. • Northern Elephant Seal-underwent an extreme population bottleneck resulting from fur hunting in the nineteeth century. • Ashkenazic Jews-a religious minority in Central Europe that has rebounded from attempted genocide. • Endangered Species
Genetic drift contributes to evolution in a number of ways • by decreasing genetic diversity, it can put the population at risk of extinction • its random nature increases the genetic differentiation between two or more populations • this may lead to speciation if one or more populations become reproductively isolated. • Genetic differentiation caused by genetic drift may change the genetic background against which new mutations act. If there is epistasis, a new mutation may be favorable in some populations and unfavorable in others. (Wright’s shifting balance theory)
The Neutral Theory of Molecular Evolution. • One of the most interesting breakthroughs in evolutionary biology in the 1960’s-1990’s, has been the development of the neutral theory of molecular evolution. • It was introduced by the Japanese theoretician Motoo Kimura, in the late 1960’s. • It is a theory of evolution that Darwin never could have anticipated (evolutionary biology does not begin and end with Darwin). • It runs in parallel with Darwinian evolution by natural selection, though its effects are most noticeable and easiest to understand on loci for which there are no differences in fitness between alleles (thus, it is called the neutral theory). • It causes change over vast spans of time, at a more-or-less constant rate, when averaged over many loci. • For that reason, it can be used to develop a “molecular clock”..to tell how long it has been since two lineages have diverged.