Download

1 / 28
280 likes | 436 Views
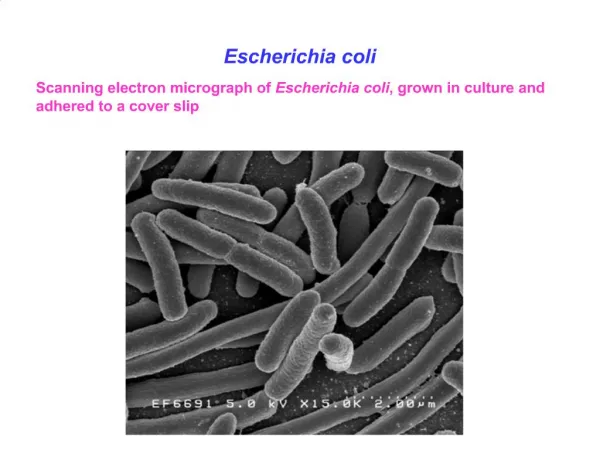
Non-random segregation of sister chromosomes in Escherichia coli. Nature 455 1248(2008). L R. E. coli chromosome organization and segregation. --chromosome territories--. 85% 15%. L R L R.

E N D
Non-random segregation of sister chromosomes in Escherichia coli Nature455 1248(2008)
L R E. coli chromosome organization and segregation --chromosome territories-- 85% 15% L R L R Replication fork-dependent locus positions are imprinted. In most generations, the <Left-Right> chromosome orientation in a mother cell is recreated as a <Left-Right-Left-Right> arrangement of sister chromosomes in daughter cells Genes. Dev. 20, 1727-1731 (2006)
--Leading strands segregate to cell poles?-- Two replichores with opposite replication polarity replisome 400kb (~56um) enzymes acting on DNA like RNA polymerases, topoisomerasesand activities like replication, repair, and recombination act as potential motors that are able to move chromosomal regions during the course of the cell cycle. Transcription direction J. Struct. Biol. 156, 273-283 (2006)
--Replichore separation-- The two arms of chromosome flank the transverse cellular axis. The left and right arms of chromosome are oriented in parallel to the long axis of the cell. J. Struct. Biol. 156, 273-283 (2006) Nat. Rev. Microbiol. 6, 28-40 (2008)
Experimental design. --Combining FROS, sbcDC and recA-- kb
Experimental design. SbcCD cleaves a hairpin formed by the palindrome on the lagging strand which is degraded in a recA- mutants by RecBCDexonuclease.
68% of the cells contain two segregated copies of the construct in the absence of induced double-strand breaks. CFP: pseudocolored green YFP: magenta Overlapping: white
Induction of SbcCD expression in a palindrome-containing strain Decrease in the number of the cells with two visibly segregated copies of the construct over time strongly associated with an increase in the number of cells with only one visible copy of the construct. Cleavage and degradation of one of the replicated chromosomes had occurred.
Cell pole bias. 89.2% 63.5% DSB doesn’t lead to cell pole migration of intact chromosome.
Cell pole bias is not observed when treated with cephalexin.
Fitting to Replication factory model? The loop constrained the lagging strands to the replisome Mol. Cell. 27, 527-538 (2007)
Model argues against the fixed replication factory model. Ovoids represent average distance traveled. oriC the replisome tracks along DNA and that the replication fork in DNA is the only determinant of replisome position. How the two independent replisomes are coordinated? L3 R3 Cell. 133, 90-102 (2008)
Concentration effect of ParA-Yfpin YLS1 • Expression level or different expression forms? • Use western to check the results • Compare the western results with those derived from plasmid stability Yfp DAPI merge DIC
Experimental design. --Plasmid Stability-- =========== RED=========== Use RED homologous recombination system to substitute the sopABC of E. coli mini-F plasmid with S. coelicolorparABS.
Experimental design. --Plasmid Stability-- Selective medium (Amp/Cm) 8hr 37 ℃ incubation 10-4 1st Non-Selective medium 12hr 37 ℃ incubation 10-4 106 dilution plating 100ul on selective/non-selective plates and count for colony number after 12hr incubation 2nd Non-Selective medium 12hr 37 ℃ incubation 10-4 3rd Non-Selective medium 12hr 37 ℃ incubation 10-4 4th Non-Selective medium 12hr 37 ℃ incubation Subculture 4 times into non-selective medium to estimate the ability of S. coelicolorparABS system in retaining the miniF plasmid in E. coli.
--Plasmid stability-- Percentage of Plasmid containing population sopABC+ sopABC- parABSP104S parABS+ # of subculture B 1 2 3 4 S. CoelicolorparABS stabilize the miniF plasmid in E. coli. P104S mutant doesn’t affect much of the system function.
Table (B2H/nucleoid localization) pT18 Cya’ • We may infer from the data: • ATP binding is crucial for nucleoid targeting as well as A-A or A-B interaction. • A-A or A-B interaction is probably independent on hydrolysis ability • --ATPase assay-- • Nucleoid targeting is independent of A-A or A-B • NC can't be masked on its N-terminus for both A-A or A-B interaction; NC could be vulnerable and suggesting the role of N-terminus in A-A interaction • --double labeling/IP-- • NC preserve some ParB interacting ability; the A-B interacting domain could lie on the C-terminus of ParB • --screen with B2H-- pT25 Cya’’
ParB-Yfp in MC1000 The foci pattern is widely observed in other parB-homolog containing organisms. Localization patterns correspond to the nucleoid Population distribution Are the localization of spots cell cycle related? Do they correspond to plasmids are other specific loci in E. coli. Yfp DAPI merge DIC 1 spot 2 spots 2 spots with one deposit 3 spots
--Alignment of ParB homology-- * ◎ * R1F ◎, R143(R80 in Spo0J), fuzzy foci (incomplete condense) with DNA binding ability/ PM R1F, presumed oligomerization with DNA binding defect/ deleted *, substituted with stop codon.
To be continued… Phenotypes of parA mutant clones in S. coelicolor Complete the control experiments in B2H Double labeling (failed) ParA/ B expression from endogenous promoter in E. coli (failed)