Download

1 / 12
330 likes | 1.03k Views
Making more plant: the biosynthesis of sucrose and starch. Biol 3470: Plant Physiol Biotechnol Lecture 9 Tues. 7 Feb. 2006 Chapter 6.1 - 6.2. From Buchanan et al., “Biochemistry and molecular biology of plants”, 2001. Making sucrose and starch is linked with photosynthesis.

E N D
Making more plant: the biosynthesis of sucrose and starch Biol 3470: Plant Physiol Biotechnol Lecture 9 Tues. 7 Feb. 2006 Chapter 6.1 - 6.2 From Buchanan et al., “Biochemistry and molecular biology of plants”, 2001
Making sucrose and starch is linked with photosynthesis • Sucrose and starch are the ultimate products of the light-independent reactions (non-recycled C) • This “fixed” carbon can be used for: • Building new plant parts by using their C skeletons directly for the biosynthesis of • Amino acids (protein) • Cell wall (polysaccharides) • Fatty acids/triglycerides • Secondary metabolites for defense (terpenes, alkaloids) • Nucleic acids (purines, pyrimidines) • ATP synthesis via respiration (glycolysis, the TCA cycle and oxidative phosphorylation)
Plant cells regulate the relative amount of carbon exported or stored in the leaf • This is known as a________ • Much of the photoassimilate is translocated immediately as sucrose in phloem • Many plants store excess fixed C in mesophyll cells to fuel mesophyll cell metabolism • starch in chloroplasts (soybean, spinach, tobacco) • sucrose in vacuole (wheat, barley, oats) • The plant must “decide” how to allocate its C from the 3-carbon carbohydrate (G3P) made in the PCR cycle • This depends on the metabolic need of sinks in the plant
Photosynthesis is the only input into plant carbon metabolism • The metabolism of carbon as carbohydrates has a central role in plant biosynthetic reactions • Photosynthesis provides the fixed carbon • Sucrose and starch are synthesized in different compartments • starch in chloroplast stroma • sucrose in mesophyll cytosol • Interchanging of this C through the hexose-phosphate pool feeds many metabolic pathways glycolysis Glyceraldehyde 3-P Photosynthesis respiration TCA cycle From Buchanan et al., “Biochemistry and molecular biology of plants”, 2001
Carbon is allocated from the hexose-P pool according to metabolic need Transport or long term C storage (in root, storage organ) More plant hv + CO2 From Buchanan et al., “Biochemistry and molecular biology of plants”, 2001 Hexose-P pool Let’s focus on the inputs and outputs of the hexose-P pool • Note how C flows from “source” (the light-independent reactions or PCR cycle) to multiple C “sinks” (starch, sucrose, C skeleton generation for making new molecules, respiration) • Cells in different stages of their life cycle require different sources of C • Rapidly dividing cells need PPP nucleic acid precursors and energy • Biosynthesizing cells need NADPH • Etc. Photosynthesis Glyceraldehyde 3-P Long (root, storage organ) or short term (leaf) C storage Energy for biosynthesis (NADPH), nucleic acid precursor (ribose 5-P) C skeletons, ATP
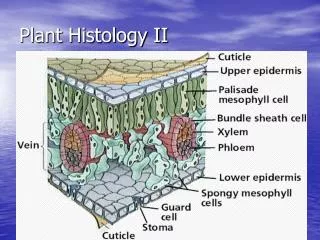
Starch and sucrose biosynthesis occur in separate cellular compartments • There are separate hexose-P pools in the stroma and mesophyll cell cytosol • Interchange between these pools occurs via a triose-phosphate translocator in the chloroplast membrane Starch is synthesized in the chloroplast stroma • Large starch grains are evident in electron micrographs of chloroplasts • Starch synthesis requires withdrawal of hexose-P from the mesophyll cellular pool • Using hexose-P for starch synthesis requires its a_________ by ATP • Only occurs in actively photosynthesizing cells Fig. 6.2 Starch grain Grana (stacked thylakoid membranes) http://www.sju.edu/biology/ksweb/microsc/mgraphs03.html
There are numerous control points that regulate the allocation of carbon • Specialized plastids called a________ store massive amounts of starch long-term • These plastids can also import and export hexose-P via a special translocator • This reflects their changing/dynamic role as a sink during seed development and source during germination • High [hexose-P]: favors starch synthesis during seed development • High inorganic P levels (substrate for starch degrading enzymes): favors starch breakdown during seed germination • The hexose-P transporter controls source/sink status of amyloplast • Note the large number of potential control points for regulating the flow of C between starch and sucrose! • Each is controlled by one or more enzymes (inside chloroplast) _____? (outside chloroplast) hexose-P pool Triose-P transporters triose-P pool Hexose-P transporter hexose-P pool triose-P pool _____? photosynthesis From Buchanan et al., “Biochemistry and molecular biology of plants”, 2001
Sucrose is synthesized in the cell cytosol from its separate hexose-P pool _________ ic hexose-P pool Glucose 1-P • Unlike starch, sucrose is synthesized in the cytosol of mesophyll cells • a/k/a photoassimilate: a disaccharide of glucose + fructose • Recall that sucrose may be • Stored • Translocated to sinks • Converted to starch via the chloroplast hexose-P pool • Metabolized via respiration to generate C skeletons and ATP • Main synthesis pathway uses hexose-P pool substrates: • Vast majority of sucrose is synthesized by sucrose phosphate synthase (SPS) and sucrose phosphate phosphatase (SPP) • Sucrose synthesis is energetically favored and allows its accumulation to high levels UTP Pi Highly energetically favored ∆G = -13 kJ / mol From Buchanan et al., “Biochemistry and molecular biology of plants”, 2001
Starch or sucrose? • How does the plant “choose” whether to make sucrose or starch? • Traditionally, it was thought that carbohydrate metabolism was governed by source-sink relationships – sink removal causes backup of starch in leaves This is false! • We now know that starch is needed in leaves for metabolic needs of the plant during dark period • Plants allocate photoassimilate between sucrose and starch in ways unrelated to: • Sink capacity of storage organs • Metabolic capability of chloroplasts in leaves to synthesize starch • Control of carbon distribution needed • Need to balance photoassimilate supply and utilization to avoid: • Depleting G3P pool in chloroplasts: keep the ____ cycle spinning! • Decreasing RuBP regeneration • Impairing carbon fixation and growth
Sucrose biosynthesis is regulated by the gluconeogenic enzyme fructose 1,6-bisphosphatase (FBPase) • The activity of the cytosolic enzyme FBPase appears to control sucrose biosynthesis • It is the first irreversible reaction in conversion of triose-P to hexose-P • Observed that • tissues with higher FBPase activity accumulate more sucrose • tissues with lower FBPase activity accumulate less sucrose From Buchanan et al., “Biochemistry and molecular biology of plants”, 2001 Translocated photoassimilate from chloroplast
The activity of FBPase is inhibited by the regulatory metabolite fructose 2,6-bisphosphate Export to chloroplast for starch biosynthesis 1 Here’s how this regulation works: • A high sucrose synthesis rate results in high cytosolic Pi levels • This in turn slows the sucrose export rate • The hexose-P pool size grows as its utilization slows • This favors Fru 2,6-P2 synthesis • High Fru 2,6-P2 inhibits FBPase from adding more to the hexose-P pool • This slows sucrose biosynthesis • The hexose-P is converted to triose-P • High triose-P levels favor the breakdown of Fru 2,6-P2, returning C to the hexose-P pool • This releases FBPase from inhibition and once again favors sucrose biosynthesis 2 3 4 3 4 2 1
The size of the hexose-P pool also directly controls the rate of photosynthesis Note that inhibition of FBPase also favors the export of triose-P to the chloroplast! • This favors starch over sucrose biosynthesis • Increasing the sizes of the hexose-P pools also feedback inhibits both the light-dependent and -independent reactions of photosynthesis • Less Pi available to make ATP in the photosynthetic E.T.C. • This in turn reduces O2 evolution and CO2 fixation • The balance between allocation to starch or sucrose synthesis is delicate and very dynamically controlled millisecond to millisecond • Controlled by compartmental levels of triose-P and Pi • Part of a complicated signal transduction network between chloroplast and cytosol • Dual role • Provide energy for growth and C-skeletons • Exchange information about metabolic status cytosol chloroplast