Download

1 / 24
240 likes | 270 Views
This theoretical model proposes a stochastic process to calculate the binding of DnaA ATP molecules to oriC, influencing DNA replication initiation in E. coli. The model considers factors like DnaA ATP binding affinity, dissociation constants, and hemimethylation effects on replication timing. It uses Monte Carlo simulations to determine binding probabilities and regulates gene site formation based on chromosome elongation dynamics, offering insights into the molecular mechanisms governing DNA replication initiation in Escherichia coli.

E N D
A model for the Initiation of Replication in Escherichia coli • Author Joseph M. Mahaffy • Judith W Zyskind J. theor. Biol. 1989
Main contributions • The number of DnaA protein monomers per origin at the time of initiation varies directly with growth rate, eliminating the possiblity that the total concentration of DnaA is the sole determinant of when initiation occurs.
DnaA states • Activator. DnaA when complexed with ATP can be bound to oriC where it serves as an activator for initiation of replication. • DnaA ATP bound form. It can be bound to any number of gene sites along the chromosome. These sites act as a reserve for DnaA ATP. • Autorepressor. DnaA protein acts as a repressor for the expression of its own gene. • DnaA ATP (free), is free in the cytoplasm to bind to any of the above gene locations including oriC • DnaA (inactive). DnaA unbound to a nucleotide, Dna ADP, and DnaA bound to acidic phospholipids.
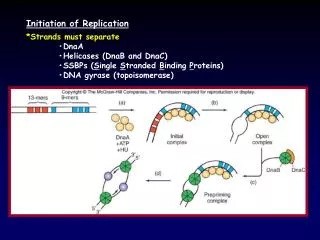
Binding to OriC • A stochastic process is used to compute the amount of DnaA ATP bound to oriC(at each t) • The model assumes that DnaA ATP molecules bind to the four binding sites in oriC with equally likely chances. • Additionally DnaA protein binds to itself to form large spherical complexes containing OriC DNA and 20-40 monomers of Dna Proteins. • In the model, additional molecules are added at the four sites with equally likely random events until an initiation event occurs. • An average rate of binding k0 is assumed • The model ignores the probable differences in binding constants and changes in the geometry as multiple molecules bind to oriC
Binding to oriC • The model also considers the possibility that a molecule of DnaA ATP may dissociate over a small interval of time with dissociation constant, kd. • The model assumes that only the upper most molecules in the complex are subject to dissociation. • Max of 4 molecules corresponding to the four binding sites are allowed to dissociated.
Hemimethylation • Before examining the stochastic binding, we need to know how long it has been since the last initiation event. • After replication begins there is an 8-10 min period during which the chromosome is bound to the membrane. • This together with the loss of supercoiling affects the binding ability of DnaA ATP. • Therefore if the last initiation event for a particular oriC has occurred in less than 8 min, the model does not allow binding.
Probabilities • The probabilistic part of the model uses a Monte Carlo simulation scheme to determine the individual binding events of Dna ATP to oriC. • Choose J so that only one binding event can occur during the time interval t /J
Probabilties • The model also allows for dissociation of at most one molecule over the interval t. • Let the probability that a DnaA ATP monomer becomes unbound be: • Kd is the kinetic constant for the reverse reaction to the binding of DnaA ATP to oriC. • N is the number of exposed molecules(<=4)
Deterministic Phase • The mathematical model assumes that there is a uniform distribution of the consensus sequence along the chromosome where DnaA ATP can bind(reserve active form). • The model adds bindings sites regularly to each elongating chromosome. • The number of actively elongating chromosomes is determined by the number of oriC’s which have an age between 0 and C, where C is the time required for replication for replication of the chromosome.(C=45min)
Deterministic Phase • The equation for the number of gene sites is: • GS(t + t) = GS(t) + AC kgs t • AC corresponds to the number of oriC’s with an age between O and C • Kgs is the rate of formation of gene sites per elongating chromosome.
Determining [DnaAg] • The equation for the concentration of DnaAg, is derived by applying mass balance laws for the formation and release of DnaA ATP monomers bound to the consensus sequences along the chromosome and including a term to account for dilution. The model does not consider the low intrinsic activity that might convert part of the bound form to DnaA ADP.
[DnaAg] Kg1 is the rate of formation of bonds between DnaA ATP in the cytoplasm with unbound gene sites, kg2 is the rate of dissociation of the bound form, and = ln (2) / is the dilution rate with the cell doubling time.
DnaA ATP in the cytoplasm [DnaAf] • The third state of DnaA in the mathematical model is that of an auto repressor for the dnaA gene. We assume that the repressor concentration is determined from the concentration of DnaA ATP monomers in the cytoplasm and from certain forms of DnaA that are in the inactive state.
[DnaAf] The first expression on the right hand side of the equation represents the production of DnaAf using standard biochemical kinetics for end product repression. The repressor concentration is determined from the concentration of DnaAf and from certain forma of DnaA that are in the inactive state. For modeling purposes we assume that the amount of DnaAi which is available for repression is a fraction of the total DnaAi.
The next step • Whenever 30 or more molecules of DnaA ATP binds to a particular oriC, an initiation event is assumed to occur. • When initiation occurs the DnaA ATP molecules are released into the cytoplasm, increasing the [DnaAf]. • A new oriC is created and both enter a hemimethylated state for 8 min. • After testing for an initiation event, the mathematical model checks for a cell division. • In order for a cell to divide, there must be two oriC’s which have an age greater than C+D, where the time required for elongation is assumed to be 45min and the period of latency prior to cell division is taken to be 25 min.
Cell Division • When the cell devides the volume is split exactly in half. • The concentrations are maintained which means that the number of molecules of all the relevant biochemical species are halved. • The amount of DnaA ATP bound to each oriC before division differs because of the stochastic process. • The next final step in the model is to increase the time by 0.1 min and repeat the process unless the termination time is reached.