Download
1 / 28
280 likes | 594 Views
第 2 节 酶的结构与功能. 1. 酶的组成成分. 一 . 酶的结构. 根据组成成分,酶可分为两类:. 单纯酶 —— 仅由蛋白质组成的酶。. 结合酶 —— 除蛋白质外,还有非蛋白质成分。 全酶 = 酶蛋白 + 辅因子. 辅因子有两种 :. 辅酶 —— 与酶蛋白结合较松弛的 小分子有机物。. 辅基 —— 与酶蛋白结合较紧密,常常以 共价键 结合,包括 小分子有机物及金属离子。. 2. 酶的聚合状态. 一 . 酶的结构. 根据酶的聚合状态,酶可分为三类:. 单体酶 —— 酶蛋白仅有一条多肽链组成。

E N D
1. 酶的组成成分 一. 酶的结构 根据组成成分,酶可分为两类: 单纯酶——仅由蛋白质组成的酶。 结合酶——除蛋白质外,还有非蛋白质成分。 全酶 = 酶蛋白 + 辅因子 辅因子有两种: 辅酶——与酶蛋白结合较松弛的小分子有机物。 辅基——与酶蛋白结合较紧密,常常以共价键结合,包括小分子有机物及金属离子。
2. 酶的聚合状态 一. 酶的结构 根据酶的聚合状态,酶可分为三类: 单体酶——酶蛋白仅有一条多肽链组成。 寡聚酶——酶蛋白是寡聚蛋白质,由几个至十多个亚基组成,以非共价键连接。 多酶复合体——由几个酶聚合而成的复合体。一般由在系列反应中功能相关的酶组成,有利于一系列反应的连续进行。
3. 同工酶 一. 酶的结构 同工酶的定义: 指具有不同分子结构但催化相同反应的一组酶。 每组同工酶中各种酶的异同: 相同点:催化相同的化学反应,大多数是寡聚酶。 不同点:体外:理化性质 体内:催化特性、分布的部位、生物学功能
一.酶的结构 3. 同工酶 以乳酸脱氢酶为例 乳酸脱氢酶 (lactate dehydrogenase,LDH)
3. 同工酶 一. 酶的结构 LDH是1959年发现的第一个同工酶。 由4个亚基组成的寡聚酶,亚基分为M型和H型。 因此可以装配成五种四聚体: H4(LDH1)、H3M(LDH2)、H2M2(LDH3)、HM3(LDH4、M4(LDH5) 不同的LDH分布在不同组织中。例如,脊椎动物心脏中主要是 LDH1,而骨骼肌的则是LDH5。
3. 同工酶 一. 酶的结构 同工酶的作用: 对于适应不同的组织、器官的不同生理需要非常重要;是代谢调节的一种重要方式。 同工酶物理性质差异: • Aa组成和顺序不同 • 催化特性不同 • 电泳行为不同 • 组织、器官中分布不同 • 生理功能不同
3. 同工酶 一. 酶的结构 不同组织中的LDH同工酶的电泳图谱
一. 酶的结构 Plant Cell Physiology, 2005.
3. 同工酶 一. 酶的结构 研究同工酶的意义: 是研究代谢调节、个体发育、细胞分化、分子遗传等方面的有力工具。 研究蛋白质结构和功能的好材料。 在临床医学、农业遗传育种、病理分析上都有应用价值 。
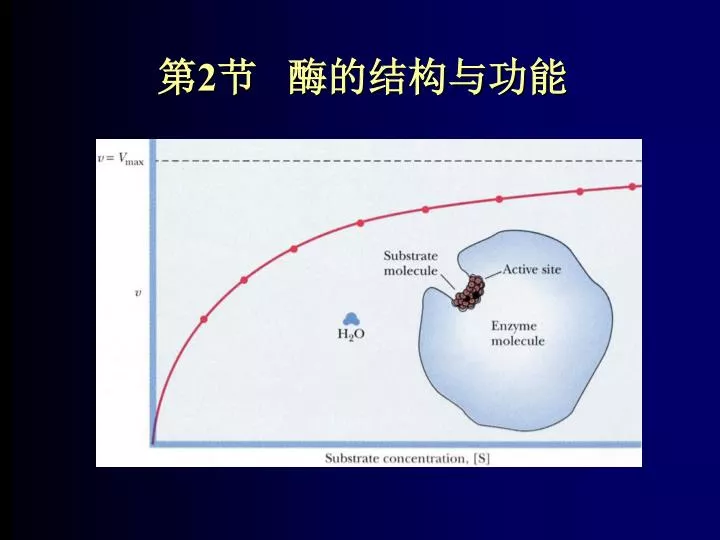
1. 酶的活性中心 二. 酶的结构与功能 由一些氨基酸残基的侧链基团组成。 对于结合酶,辅因子常常是活性中心的组成部分。 指酶分子中直接和底物结合,并和酶催化作用直接有关的部位。 (1)酶活性中心的组成: 这些基团在一级结构上可能相距很远,甚至可能不在一条肽链上,但在蛋白质空间结构上彼此靠近,形成具有一定空间结构的区域。
1. 酶的活性中心 二. 酶的结构与功能
1. 酶的活性中心 二. 酶的结构与功能 The structure of a glycogen phosphorylase monomer
1. 酶的活性中心 二. 酶的结构与功能 (2)酶活性中心的特点 Substrates typically lose waters (of hydration 水合作用) in the formation of the ES complex
1. 酶的活性中心 二. 酶的结构与功能 (2)酶活性中心的特点 1. 活性中心在酶分子总体积中只占相当小的部分 (约1%2%),相当于23个氨基酸残基。 2. 都是酶分子表面的一个凹穴,有一定的大小和形状,但不是刚性的,而具有一定的柔性。 3. 活性中心为非极性的微环境,有利于与底物结合。
1. 酶的活性中心 二. 酶的结构与功能 (2)酶活性中心的特点 4. 底物与酶通过形成较弱键力的次级键相互作用并结合到酶的活性中心。 5. 酶的活性部位并不是和底物的几何图形正好吻合,而是在酶与底物结合的过程中,底物分子或酶分子或它们两者的构象同时发生一定变化后才相互契合,这时催化基团的位置也正好处于所催化底物的敏感化学键部位。
1. 酶的活性中心 二. 酶的结构与功能 结合部位:底物在此与酶分子结合。一个酶的结合部位又可以分为各种亚位点,分别与底物的不同部位结合。 催化部位:底物的敏感键在此被打断或形成新的键,从而发生一定的化学反应。一个酶的催化部位可以不止一个。 (2)有关酶活性中心的几个术语
1. 酶的活性中心 二. 酶的结构与功能 有些酶的分子表面除了活性中心外,还具有重要的功能部位——调节中心 调节中心可以与小分子的代谢物相结合,使酶分子的构象发生改变,从而影响酶的活性。这种作用叫变构效应(又叫别构效应); 具有变构效应的酶叫变构酶,引起变构的小分子物质叫变构剂(调节物)。
二. 酶的结构与功能 使酶活性升高的变构叫正变构,此时的变构剂叫正变构剂(正调节物); 使酶活性降低的变构叫负变构,此时的变构剂叫负变构剂(负调节物)。
2. 变构酶 二. 酶的结构与功能 变构酶(别构酶)是指一些含有2个或2个以上亚基的寡聚酶,在变构酶分子上,别构效应剂的调节部位一般远离活性中心,但活性部位与调节部位之间或者活性部位之间,存在着相互作用(变构效应,协同效应)。调节物与酶分子的调节部位结合之后,引起酶分子构象发生变化,从而提高或降低活性部位的酶活性
2. 变构酶 二. 酶的结构与功能 已知的变构酶都是寡聚酶。 变构酶分子上除了活性中心外,还有调节中心。这两个中心处在酶蛋白的不同部位,有的在不同的亚基上,有的在同一亚基上。 变构酶的 v-[S] 的关系不符合米氏方程,所以其曲线不是双曲线型。 变构酶的特点:
2. 变构酶 二. 酶的结构与功能 别构酶动力学曲线 A.为非调节酶的曲线B.为别构酶的S形曲线
二. 酶的结构与功能 2. 变构酶 完整的酶分子 催化亚基 调节亚基 (活性形式) (三聚体) (二聚体) E.coli ATCase(天冬氨酸转氨甲酰酶)中亚基排列及全酶与亚基的关系
二. 酶的结构与功能 2. 变构酶
二. 酶的结构与功能 3. 诱导酶 诱导酶(inducenzyme)是细胞内在正常状态下一类很少存在或没有的酶,当细胞中因加入了诱导物后而被诱导产生的酶,它的含量在诱导物存在下显著增高,这种诱导物往往是该酶底物的类似物或底物本身。 诱导酶是20世纪四十年代,微生物学工作者在研究大肠杆菌以葡萄糖和山梨糖醇为培养基时出现二度生长时发现的.
二. 酶的结构与功能 3. 诱导酶 即第一次生长的量与葡萄糖成比例,第二次生长的量与山梨糖醇浓度成比例。二度生长现象是由于一般大肠杆菌细胞内只有利用葡萄糖的酶,而不含利用山梨糖醇的酶。因此对葡萄糖的利用不需要适应,即可利用,而对山梨糖醇的利用必需在山梨糖醇的诱导下,生成能分解山梨糖醇的酶才能进行,但这种诱导作用受到葡萄糖的阻遏,故必须在葡萄糖消耗完后才逐渐利用山梨糖醇。
二. 酶的结构与功能 3. 诱导酶 E.coli二度生长现象 a:葡萄糖50 μg/mL,山梨糖醇150 μg/mL;b:葡萄糖100 μg/mL,山梨糖醇100μg/mL;C:葡萄糖150μg/mL;山梨糖醇50 μg/mL)
