Download
1 / 1
10 likes | 268 Views
Cell cycle synchronization in male and female gametes of tobacco ( Nicotiana tabacum ). Introduction

E N D
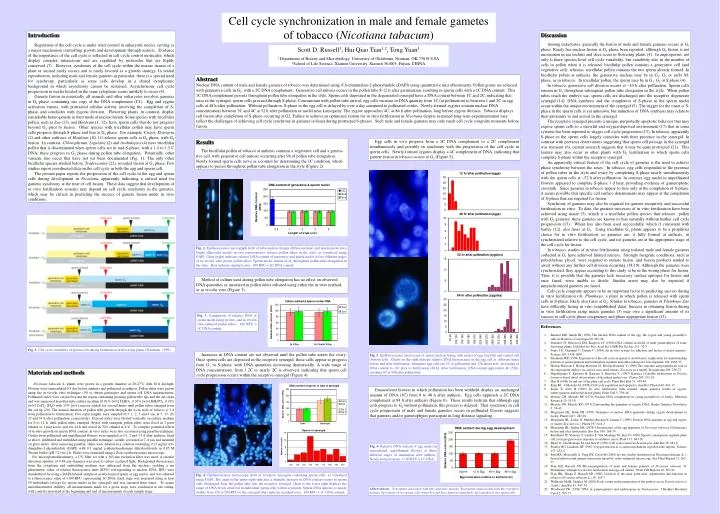
Cell cycle synchronization in male and female gametes of tobacco (Nicotiana tabacum) Introduction Regulation of the cell cycle is under strict control in eukaryotic nuclei, serving as a major mechanism controlling growth and development through mitosis. Evidence of the importance of the cell cycle is reflected in cell cycle control molecules, which display complex interactions and are regulated by molecules that are highly conserved (7). However, synchrony of the cell cycle within the mature tissues of a plant or animal rarely occurs and is rarely favored as a growth strategy. In sexual reproduction, including male and female gametes in particular, there is a special need for synchrony, particularly as some cells develop in a shared cytoplasmic background in which asynchrony cannot be sustained. Asynchronous cell cycle progression in nuclei located in the same cytoplasm seems unlikely to occur (4). Gamete fusion in essentially all animals and other eukaryotes involves gametes in G1 phase, containing one copy of the DNA complement (C1). Egg and zygote activation ensues, with protracted cellular activity involving the completion of S-phase, and concludes with zygotic division. Flowering plants, in contrast, display remarkableheterogeneityintheirmodeofnuclearfusion.Somespecieswith tricellular pollen,suchasZea(13),andHordeum(1, 12), have sperm cells that do not progress beyond G1 prior to fusion. Other species with tricellular pollen may have sperm cells progress through S-phase and fuse at G2 phase. For example, Crepis, Elytrigria (2) and other cultivars of Hordeum (10, 11) release sperm cells at G2 phase, with G2 fusion. In contrast, Chlorophytum, Ligularia (2) and Arabidopsis (4) have tricellular pollen that is disseminated when sperm cells are in mid-S-phase, with a 1.4 to 1.5 C DNA; these progress to G2 phase during pollen tube elongation. Potentially, other variants also occur that have not yet been documented (Fig. 1). The only other bicellular species studied before, Tradescantia (22), revealed fusion at G1 phase. Few studies report coordinated data on the cell cycle in both the egg and sperm cell. The present paper reports the progression of the cell cycle in the egg and sperm cells during development in Nicotiana, apparently indicating a critical need for gametic synchrony at the time of cell fusion. These data suggest that development of in vitro fertilization systems may depend on cell cycle synchrony in the gametes, which may be critical in predicting the success of gamete fusion under in vitro conditions. Discussion Among eukaryotes, generally the fusion of male and female gametes occurs at G1 phase. Rarely has nuclear fusion at G2 phase been reported, although G2 fusion is not uncommon in sea urchins and does occur in flowering plants (4). In angiosperms, not only is there species-level cell cycle variability, but variability also in the number of cells in pollen when it is released: bicellular pollen contains a generative cell (and vegetative cell), whereas tricellular pollen contains the two sperm already formed. In bicellular pollen at anthesis, the generative nucleus may be in G1, G2 or early M-phase, as in tobacco. In tricellular pollen, the sperm may be in G1, G2 or S phase (4). In tobacco, generative cell division occurs at ~10 h after pollination. Sperm cells remain in G1 throughout subsequent pollen tube elongation in the style. When pollen tubes reach the embryo sac, sperm cells are discharged into the receptive degenerate synergid(14). DNA synthesis and the completion of S-phase in the sperm nuclei occurwithintheuniqueenvironmentofthesynergid(5).Thetriggerfortheonset of S-phaseinthespermcellsisyetunknown,butinductionof DNA synthesismayrelatetotheirproximity to and arrival in the synergid. The receptive synergid presents a unique, purportedly apoptotic behavior that may expose sperm cells to a stressful and oxygen-deprived environment (17) that in some systems has been reported to trigger cell cycle progression(15).Intobacco, apparently S-phase in the sperm cells largely coincides with their presence in the synergid. In contrast with previous observations suggesting that sperm cell passage in the synergid was transient (6), current research suggests that it may be quite protracted (21). This featuremayalsooccurinotherplantswithG2fertilizationinwhichspermcells complete S-phase within the receptive synergid. An apparently critical feature of the cell cycle of gametes is the need to achieve phase synchrony between the sexes. In tobacco, egg cells responded to the presence of pollen tubes in the style and ovary by completing S-phase nearly simultaneously with thespermcellsat~52hafterpollination.Incontrast,eggnucleiinunpollinated flowers appeared to complete S-phase 1 d later, providing evidence of gametophytic crosstalk. Since gametes in tobacco appear to fuse only at the completion of S-phase, it seems possible that specific cell surface determinants may appear at the completion of S-phase that are required for fusion. Synchrony of gametes may also be required for gamete receptivity and successful fertilization in vitro. To date, the greatest successes of in vitro fertilization have been achieved using maize (3), which is a tricellular pollen species that releases pollen with G1 gametes; these gametes are known to fuse naturally without further cell cycle progression (13). Wheat has also been used successfully, which if consistent with barley (12), also fuses at G1. Using tricellular G1 plants appears to be a propitious choice for in vitro fertilization, as gametes are: i) fully formed at anthesis, ii) synchronized relative to the cell cycle, and iii) gametes are at the appropriate stage of the cell cycle for fusion. In tobacco, studies of in vitro fertilization using isolated male and female gametes collected at G1 have achieved limited success. Strongly fusigenic conditions, such as polyethylene glycol, were required to initiate fusion, and fusion products tended to arrest without any further cell division occurring (18,19). Although the gametes were synchronized, they appear according to this study to be in the wrong phase for fusion. Thus, it is possible that the gametes lack necessary surface epitopes for fusion and once fused, were unable to divide. Similar arrest may also be expected if unsynchronized gametes are fused. Cell cycle congruity appears to be an important factor in predicting success during in vitro fertilization (4). Plumbago, a plant in which pollen is released with sperm cells in S-phase, likely also fuses at G2. Similar to tobacco, gametes of Plumbago also have difficulty fusing in vitro (unpublished data). Success in obtaining fusion during in vitro fertilization using maize gametes (3) may owe a significant amount of its success to cell cycle phase congruency and phase appropriate fusion (13). Scott D. Russell1, Hui Qiao Tian1,2, Tong Yuan1 1Department of Botany and Microbiology, University of Oklahoma, Norman, OK 73019 USA 2School of Life Science, Xiamen University, Xiamen 361005, Fujian, CHINA Abstract Nuclear DNA content of male and female gametes of tobacco was determined using 4',6-diamindino-2-phenylindole (DAPI) using quantitative microfluorimetry. Pollen grains are released with generative cells in G2, with a 2C DNA complement. Generative cell mitosis occurs in the pollen tube 8-12 h after germination, resulting in sperm cells with a 1C DNA content. This 1C DNA complement persists throughout pollen tube elongation in the style. Sperm cells deposited in the degenerated synergid have a DNA content between 1C and 2C, indicating that once in the synergid, sperm cells proceed through S-phase. Concomitant with pollen tube arrival, egg cells increase in DNA quantity from 1C (at pollination) to between 1 and 2C in egg cells at 48 h after pollination. Without pollination, S-phase in the egg cell is delayed by over a day compared to pollinated ovules. Newly-formed zygotes contain nuclear DNA concentrations between 3C and 4C at 52 h after pollination as nuclei near karyogamy. The zygote approaches 4C, at 84 h after pollination, long before zygote division. Tobacco displays cell fusion after completion of S-phase occurring at G2. Failure to achieve an optimized system for in vitro fertilization in Nicotiana despite sustained long-term experimentation may reflect the challenges of achieving cell cycle synchrony in gametes isolated during protracted S-phases. Such male and female gametes may only reach cell cycle congruity moments before fusion. Results The bicellular pollen of tobacco at anthesis contains a vegetative cell and a genera-tive cell, with generative cell mitosis occurring after 9h of pollen tube elongation. Newly-formed sperm cells serve as a control for determining the 1C condition, which appears to persist throughout pollen tube elongation in the style (Figure 2). Egg cells in vivo progress from a 1C DNA complement to a 2C complement simultaneously and possibly in synchrony with the progression of the cell cycle in sperm cells. Newly-formed zygotes display a 4C complement of DNA, indicating that gamete fusion in tobacco occurs at G2 (Figure 5). Fig. 2. Epifluorescence micrograph (left) of tube nucleus (longer diffuse nucleus) and sperm nuclei (two bright ellipsoidal nuclei) in two representative tobacco pollen tubes in the style, as visualized using DAPI. Chart (right) indicates relative DNA content of generative and sperm nuclei at five different stages of in vivo/in vitro grown pollen tubes. Sperm nuclei remain in G1 throughout pollen tube elongation in the style. Bars indicate standard error. 100 RFU 1C DNA content. Method of culture used during pollen tube elongation has no effect on observed DNA quantities as measured in pollen tubes cultured using either the in vitro method, or in vivo/in vitro (Figure 3). Fig. 3. Comparison of relative DNA of spermnucleiusinginvitro-andin vivo/in vitro-cultured pollen tubes. 100 RFU 1C DNA content. • References • 1. Bennett MD, Smith JB (1976) The nuclear DNA content of the egg, the zygote and young proembryo cells in Hordeum. Caryologia 29: 435-46 • 2. Ermakov IP, Morozova EM, Karpova LV (1980) DNA content in nuclei of male gametophytes of some flowering plants. Dokl Bot Sci Proc Acad Sci USSR Bot Sci Sec 251: 32-3 • 3. Faure J-E, Digonnet C, Dumas C (1994) An in vitro system for adhesion and fusion of maize gametes. Science 263: 1598-1600 • 4. Friedman WE (1999) Expression of the cell cycle in sperm of Arabidopsis: implication for understanding patterns of gametogenesis and fertilization in plants and other eukaryotes. Development 126: 1065-75 • 5. Heslop-Harrison J, Heslop-Harrison JS, Heslop-Harrison Y (1999) The structure and prophylactic role of the angiosperm embryo sac and its associated tissues: Zea mays as a model. Protoplasma 209: 256-72 • 6. Higashiyama T, Kuroiwa H, Kawano S, Kuroiwa T. (1997) Kinetics of double fertilization in Torenia fournieri based direct observations of the naked embryo sac. Planta 203: 101-10 • 7. Hirt H (1996) In and out of the plant cell cycle. Plant Mol Biol 31: 459-64 • 8. King KL, Cidlowski JA (1998) Cell cycle regulation and apoptosis. Ann Rev Physiol 60: 601-17 • 9. Kranz E, Lörz H (1993) In vitro fertilization with isolated, single gametes results in zygotic embryogenesis and fertile maize plants. Plant Cell 5: 739-46 • 10. Mericle LW, Mericle RP (1970) Nuclear DNA complement in young proembryos of barley. Mutation Research 10: 515-8 • 11. Mericle LW, Mericle RP (1973) Confounding the quandary of zygotic DNA. Barley Genetics Newsletter 3: 39-42 • 12. Mogensen HL, Holm PB (1995) Dynamics of nuclear DNA quantities during zygote development in barley. Plant Cell 7: 487-94 • 13. Mogensen HL, Leduc N, Matthys-Rochon E, Dumas C (1995) Nuclear DNA amounts in egg and zygote of maize (Zea mays L.). Planta 197: 641-5 • 14. Mogensen HL, Suthar HK (1978) Ultrastructure of the egg apparatus of Nicotiana tabacum (Solanaceae) before and after fertilization. Bot Gaz 140: 168-79 • 15. Reichheld JP, Vernoux T, Lardon F, Van Montagu M, Inzé D (1999) Specific checkpoints regulate plant cell cycle progression in response to oxidative stress. Plant J 17: 647-56 • 16. Shaul O, Van Montagu M, and Inze D (1996) Cell cycle control in Arabidopsis. Ann Bot 78: 283-8 • 17. Stanley RG, Linskens HF (1967) Oxygen tension as a control mechanism in pollen tube rupture. Science 157: 833-4 • 18. Sun MX, Moscatelli A, Yang HY, Cresti M (2000) In vitro double fertilization in Nicotiana tabacum (L.): fusion behavior and gamete interaction traced by video-enhanced microscopy. Sex Plant Reprod 12: 267-75 • 19. Tian HQ, Russell SD Micromanipulation of male and female gametes of Nicotiana tabacum: II. Preliminary attempts for in vitro fertilization and egg cell culture. Plant Cell Reports 16: 657-61 • 20. Tian HQ, Zhang Z, Russell SD (1998) Isolation of the male germ unit: organization and function in tobacco (Nicotiana tabacum L.) 18: 143-7 • 21. Wallwork MAB, Sedgley M (2000) Early events in the penetration of the embryo sac in Torenia fournieri (Lind.). Ann Bot 85: 447-54 • 22. Woodward JW (1956) DNA in gametogenesis and embryogeny in Tradescantia. J Biophys Biochem Cytol 2: 765-77 Fig. 1. Cell cycle variability of sperm cells during fertilization in flowering plants (Friedman, 1999). Increases in DNA content are not observed until the pollen tube enters the ovary. Once sperm cells are deposited in the receptive synergid, these cells appear to progress from G1 to S-phase, with DNA quantities increasing dramatically. A wide range of DNA concentrations, from 1.2C to nearly 2C is observed, indicating that sperm cell cycle progression occurs within the receptive synergid (Figure 4). Fig. 5. Epifluorescence microscopy of sperm nucleus fusing with nuclei of egg (top left) and central cell (bottom left). Charts on the right indicate relative DNA fluorescence in the egg cell at different times before and after fertilization. Immature egg cells are 1C at pollination and 12h afterwards, increasing in DNA content to ~2C prior to fertilization (48 h). After fertilization, DNA content approaches 4C (52h), reaching 4C at 84h after pollination. Materials and methods Nicotiana tabacum L. plants were grown in a growth chamber at 20-27C with 16 h daylight. Flowers were emasculated 0.5 day before anthesis and pollinated at anthesis. Pollen tubes were grown using the in vivo/in vitro technique (19) to obtain generative and sperm cell DNA measurements. Pollinated styles were cut just beyond the region containing growing pollen tube tips and the cut stylar end was immersed in pollen tube culture medium (0.01% [w/v] H3BO3, 0.01% [w/v] KH2PO4, 0.01% [w/v] CaCl2·2H2O with 15% [w/v] sucrose added) for several hours until pollen tubes emerged from the cut tip (20). The normal duration of pollen tube growth through the 4-cm style of tobacco is 2 d from pollination to fertilization. Five stylar lengths were sampled (0.5, 1, 2, 3 and 4 cm, at 5, 13, 20, 27 and 34 h after pollination, respectively). Excised styles were floated vertically on culture medium for 6 to 12 h, until pollen tubes emerged. Styles with emergent pollen tubes were fixed in 3 parts ethanol to 1 part acetic acid for 24 h and stored in 70% ethanol at 4C. To compare potential effects of in vitro growth on sperm DNA content, in vivo styles were also sectioned using paraffin technique. Ovules from pollinated and unpollinated flowers were sampled at 0.5, 2 and 3.5 d after anthesis, fixed asabove,infiltratedandembeddedusingparaffin technique, serially sectionedat 7–8 µm and mounted on glass slides. After removing paraffin, slides were labeled in a solution containing 0.25 µg/ml 4',6-diamidino-2-phenylindole (DAPI) with 0.1 mg/ml ρ-phenylenediamine dihydrochloride in 0.05 M Trizma buffer (pH 7.2) for 1 h. Slides were examined using a Zeiss epifluorescence microscope. For microspectrofluorimetry, a UV filter set with a 365-nm excitation filter was used. A circular detection aperture of 9.84 µm diameter was used to reduce scattered light. Background fluorescence from the cytoplasm and embedding medium was subtracted from the nucleus, yielding a net photometric value of relative fluorescence units (RFU) corresponding to nuclear DNA. RFU were standardized by using a DAPI-labeled sample of newly-formed sperm or egg nuclei, and was adjusted to a fluorescence value of 100 RFU, representing 1C DNA. Each stage was measured using at least 50 individuals (except for sperm nuclei in the synergid) and was repeated three times. To assure microfluorimeter stability, all measurements made for a given stage were conducted at one sitting, with controls measured at the beginning and end of measurements at each sample stage. Emasculatedflowersinwhichpollinationhasbeenwithheld display an unchanged amount of DNA (1C) from 0 to 48 h after anthesis. Egg cells approach a 2C DNA complement at 84 h after anthesis (Figure 6). These results indicate that although egg cells progress to G2 without pollination, this process is delayed. That synchrony in cell cycle progression of male and female gametes occurs in pollinated flowers suggests that gametes and/or gametophytes participate in long distance signaling. Fig. 6. Relative DNA content of egg nuclei in emasculated, unpollinated flowers at three different stages of maturation after anthesis. Newly-formed sperm = 100 RFU 1C DNA. Fig. 4. Epifluorescence microscopy (left) of receptive synergids containing sperm cells, as visualized using DAPI. The chart in the upper right indicates a dramatic increase in DNA content occurs in sperm cells discharged from the pollen tube into the receptive synergid. Chart to the lower right displays the range of DNA levels observed in individual sperm cells within synergids. Sperm DNA appears to nearly double from 120 to 200 RFU in the synergid. Bars indicate standard error. 100 RFU 1C DNA content. Abbreviations: Svn=sperm associated with the vegetative nucleus; Sua=sperm unassociated with the vegetative nucleus; Sp1=larger of two sperm cells (when Svn and Sua cannot be identified); Sp2=smaller of two sperm cells.