Download
1 / 67
670 likes | 1.11k Views
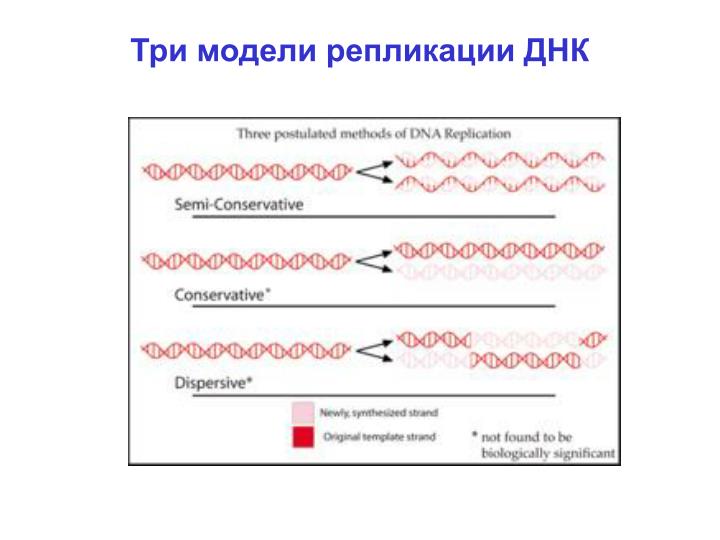
Три модели репликации ДНК. Правильная - полуконсервативная (Matthew Meselson и Franklin Stahl , 1958). Коетки растут на среде с 15 N ( тяжелый изотоп ) потом их переносят на среду с 14 N ( легкий изотоп ). Q: Что было бы для двух других моделей ?. Реплицирующаяся ДНК.

E N D
Правильная - полуконсервативная (Matthew Meselson и Franklin Stahl, 1958) Коетки растут насреде с 15N (тяжелый изотоп) потом их переносятна среду с 14N (легкий изотоп) Q: Что было бы для двух других моделей?
Проблема полярности; фрагменты Окадзаки
Центральная догма(F.Crick) ДНКРНКбелок
РНК • мРНК - перенос информации от ДНК к белку • хранение генетической информации (вирусы и фаги) • рибозимы - катализ • рРНК - структура (+ катализ) • тРНК - трансляция • регуляция
Транскрипция: инициаторный комплекс
Промоторы recA ttgatactgtat---gagcatacagtataat rpsL ttgacaccttttc--ggcatcgccctaaaat aroF ttgaaaacttta---ctttatgtgttatcgt lpdA tttaaaaattgtt--aacaattttgtaaaat cyoA tttacagtaatgt--aaccttcccgtaaaat uvrD ttggcatctctg---acctcgctgatataat atpI ttgaaatcacgg---gggcgcaccgtataat tonB ttgaatatgattgc-tatttgcatttaaaat rpsU tttacaaagcagca-gcaattgcagtaaaat cirA ttgataattgtta--tcgtttgcattatcgt fur ttttcatttaggc--gtggcaattctataat ampC ttgtcacgctga---ttggtgtcgttacaat uvrB gtgatgaactgttt-ttttatccagtataat argC ttgacacacct----ctggtcatgatagtat rpmH ttgactccggagt--gtacaattattacaat rplT ttaacgttttta---actttttaattagaat gyrA tttacctcaaactgcgcggctgtgttataat
Слабые промоторы carB ttatcgagttaattgagcagtaccgtaaaac nagB tttactggctaaa--ccagaaaacttatttt ilvG ttgccagcccacgg-tcggtcgacttactgt cysK atgtcattattt---cccttctgtatataga smp ttgcttctggca---acattaagtctcaaat amn gtgacatacta----tcggatgtgcggtaat ileR atgccagcagag---ttttttatgcgttaat fabB ttgttcggcgtac--aagtgtacgctattgt dnaN atatcaaagaag---atttttcaaatttaat argC tcgataaatggcg--gtaatttgtttttcat araB ctgacgcttttt---atcgcaactctctact sulA ttgtcactggatgt-actgtacatccataca trpR gttactgatccgc--acgtttatgatatgct dnaN ttgcaggaaaaact-ggtcaccatcgacaat cspA ttgcatcacccgc--caatgcgtggcttaat rpsU tcgcccctggaga--aagcctcgtgtatact rpoH ttcacaagcttgcattgaacttgtggataaa
Lac -промотор gcgcccaatacgcaaaccgcctctccccgcgcgttggccgattcattaatgcagctggca cgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagct cactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggaat tgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgaattcgagct cggtacccggggatcctctagagtcgacctgcaggcatgcaagcttggcactggccgtcg ttttacaacgtcgtgactgggaaaaccctggcgttacccaacttaatcgccttgcagcac atccccctttcgccagctggcgtaatagcgaagaggcccgcaccgatcgcccttcccaac agttgcgcagcctgaatggcgaatggcgcctgatgcggtattttctccttacgcatctgt
Lac -промотор cgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagct cactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggaat tgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgaattcgagct
Lac -промотор cgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagct cactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggAat tgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgaattcgagct
Lac -промотор cgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagct cactcattaggcaccccaggcTTtACActttatgcttccggctcgTATgtTgtgtggAat tgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgaattcgagct
Регуляция транскрипции у прокариот • Фактор транскрипции: взаимодействие с РНК-полимеразой • репрессоры • активаторы • Лиганд (кофактор): • способствует/препятствует связыванию с ДНК
Регуляция транскрипции trp оперона
Примеры • триптофановый репрессор • биосинтез триптофана • связан с ДНК при наличии триптофана • лактозный репрессор • катаболизм (использование) лактозы • связан с ДНК при отсутствии лактозы • арабинозный активатор • катаболизм (использование) арабинозы • связан с ДНК при наличии арабинозы
Катаболитная репрессия • CRP • активатор генов катаболизма глюкозы • репрессор генов катаболизма других сахаров • Локальные активаторы и репрессоры • арабинозный активатор • лактозный репрессор
Регуляция транскрипции у эукариот
Кроссинговер и рекомбинация • гены с одной хромосомы наследуются не независимо • рекомбинация позволяет картировать гены (и мутации)
Коллинеарность гена и белка (Brenner, 1964)
Коллинеарность гена и белка (Charles Yanofsky, 1967)
Генетический код • Генетический код – соответствие между ДНК и белком (George Gamow, 1954 (Георгий Гамов) • Крик и соавторы (1961): • не перекоывающийся (одна мутация – одна аминокислота) • вырожденный (несоклько кодонов для одной аминокислоты) • без запятых (меркеров между кодонами) • периодический
кодон - триплет • мутации, вызванные актидиновыми красителями • не протекающие (функция не ослабляется, а исчезает) • механизм – вставки/делеции оснований(дальше все путается код без запятых) CUACUACUACUACUACUACUACUACUACUACUACUACUA LeuLeuLeuLeuLeuLeuLeuLeuLeuLeuLeuLeuLeu вставка CUACUACUACGUACUACUACUACUACUACUACUACUACU LeuLeuLeuArgThrThrThrThrThrThrThrThrThr делеция CUACUACUACUACUACUACUACUACUACACUACUACUAC LeuLeuLeuLeuLeuLeuLeuLeuLeuHisTyrTyrTyr G U
Двойные мутанты и ревертанты • два класса мутаций: (+) и (–) • двойной мутант (+)¤(+) и (–)¤(–) – все еще мутантные фенотипы • двойные мутанты (+)¤(–) и (–)¤(+) – протекающие фенотипы CUACUACUACGUACUACUACUACUACUACUACUACUACU LeuLeuLeuArgThrThrThrThrThrThrThrThrThr ¤ CUACUACUACUACUACUACUACUACUACACUACUACUAC LeuLeuLeuLeuLeuLeuLeuLeuLeuHisTyrTyrTyr CUACUACUACGUACUACUACUACUACUACACUACUACUA LeuLeuLeuArgThrThrThrThrThrThrLeuLeuLeu
Тройные мутанты – реверсия! • тройные мутанты из одного класса, (+)¤(+)¤(+) и (–)¤(–)¤(–), дают протекающие фенотипы CUACUACUACGUACUACUACUACUACUACUACUACUACUACU LeuLeuLeuArgThrThrThrThrThrThrThrThrThrThr ¤ CUACUACUACUACUACUACGUACUACUACUACUACUACUACU LeuLeuLeuLeuLeuLeuArgThrThrThrThrThrThrThr двойной мутант – нет функции CUACAUCUACGUACUACUACGUACUACUACUACUACUACUAC LeuLeuLeuArgThrThrThrTyrTyrTyrTyrTyrTyrTyr ¤ CUACUACUACUACUACUACUACUACUACGUACUACUACUACU LeuLeuLeuLeuLeuLeuLeuLeuLeuArgThrThrThrThr тройной мутант – слабая функция CUACUACUACGUACUACUACGUACUACUACGUACUACUACUA LeuLeuLeuArgThrThrThrTyrTyrTyrValLeuLeuLeu
Связывание триплетов(из лекции Крика, 1966)
TTT F TCT S TAT Y TGT C TTC F TCC S TAC Y TGC C TTA L TCA S TAA stop TGA stop TTG L TCG S TAG stop TGG W CTT L CCT P CAT H CGT R CTC L CCC P CAC H CGC R CTA L CCA P CAA Q CGA R CTG L CCG P CAG Q CGG R ATT I ACT T AAT N AGT S ATC I ACC T AAC N AGC S ATA I ACA T AAA K AGA R ATG M/ start ACG T AAG K AGG R GTT V GCT A GАT D GGT G GTC V GCC A GАC D GGC G GTA V GCA A GАA E GGA G GTG V GCG A GАG E GGG G Генетический код: синонимы
