Download
1 / 39
400 likes | 556 Views
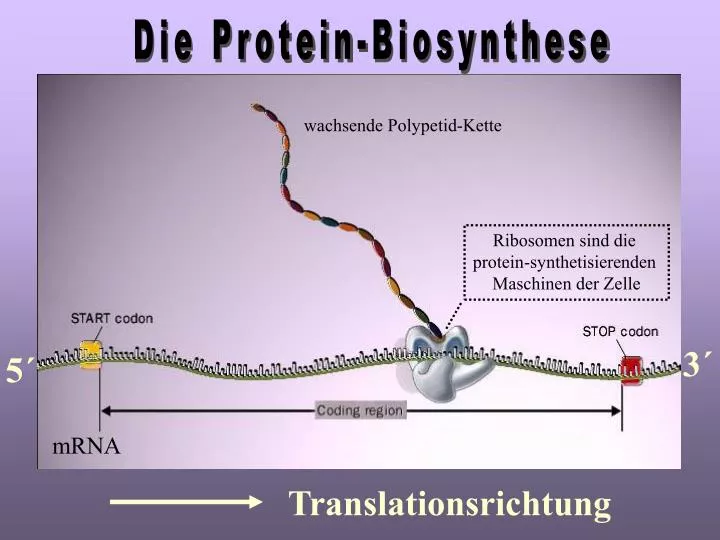
Die Protein-Biosynthese. wachsende Polypetid-Kette. Ribosomen sind die protein-synthetisierenden Maschinen der Zelle. 3´. 5´. mRNA. Translationsrichtung. Fragen aus der schriftlichen Physikumsprüfung. Aufbau der Ribosomen. 60S Untereinheit. 40S Untereinheit. 80S Ribosom.

E N D
Die Protein-Biosynthese wachsende Polypetid-Kette Ribosomen sind die protein-synthetisierenden Maschinen der Zelle 3´ 5´ mRNA Translationsrichtung
Aufbau der Ribosomen 60S Untereinheit 40S Untereinheit 80S Ribosom Jede menschliche Zelle besitzt ca. 1 Million Ribosomen (bei E. coli ca. 15 000). In Zusammenarbeit mit mRNA, tRNA und weiteren Proteinfaktoren koordinieren die Ribosomen die Proteinsynthese
Aufbau der Ribosomen Bakterien Eukaryonten 70S Ribosom 2.5 MDa 80S Ribosom 4.2 MDa 60S 50S 40S 30S 40S
Ribosomen bauen sich aus rRNA und r-Proteinen auf 16S rRNA Bakterien 70S Ribosom 2.5 MDa 50S 23S rRNA (3200 Nt) 5S rRNA (120 Nt) 34 r-Proteine 5‘ 3‘ 30S 16S rRNA (1540 Nt) 21 r-Proteine
16S rRNA Die Kristallstruktur der 30S Untereinheit 5‘ 3‘
Nukleolus Zellkern rDNA 5S rRNA Prä-rRNA (45S) Ribosomale Proteine (L, S) 28S 5.8S 18S 5S Präribosomale Partikel 28S 18S 5.8S Zytoplasma 5S 40S Untereinheit 60S Untereinheit Die Ribosomen-Biogenese läuft im Nukleolus ab >> eukaryontische Ribosomen entstehen im Zellkern, genauer im Nukleolus, und müssen von dort über das Nukleoplasma ins Zytoplasma gelangen, wo sie die mRNA translatieren Die komplizierte Ribosomen-Entstehung im Nukleolus erfordert zeitliche und räumlicheKoordination von vielen Teilprozessen. Dies wird durch die hohe strukturelle Organisation des Nukleolus gewährleistet.
Ribosomen synthetisieren Polypeptide mRNA 60S Bereich für Peptidsynthese 40S Ausgang für die Polypeptidkette diemRNA liegt wie ein Kabel auf einer Plattform in einer Einbuchtung der 40S bzw. 60S Untereinheit. Dort ist auch der Bereich der Peptid-Synthese. Die wachsende Polypeptid-Kette tritt durch einen Art Tunnel innerhalb der großen Untereinheit aus dem Ribosom heraus. Nach der Polypeptid-Synthese faltet sich die Aminosäure-Kette in ihre korrekte 3-D-Konformation
mRNA Shine-Delgarno-Sequenz Prokaryontische mRNA 3‘ 5‘ fMet 5‘ 16S rRNA 3‘ fMet fMet Eukaryontische mRNA 3‘ 5‘-Kappe m7G 50S Unter- einheit 40S Untereinheit Ribosomen-Scan 3‘ 5‘-Kappe m7G Die Initiation der Protein-Synthese InitiationsfaktorIF-3 bindet an die 30S Untereinheit, was die Anlagerung der 50S Untereinheit zunächst vehindert Anbindung der mRNA: Shine-Delgarno-Sequenz komplementär zum 3‘-Ende der 16S rRNA >> Positionierung des AUG im P-Bereich Bindung der fMethionyl-tRNA im P (=Peptidyl)-Bereich durch Codon::Anticodon-Wechselwirkung. Die tRNAfMet kann nur im P-Bereich, nicht im A-Bereich binden, was durch IF2 kontrolliert wird. IF2 ist eine GTPase. Unter GTP-Hydrolyse durch IF2 kann schließlich die 50S Untereinheit andocken, wobei IF2 und IF3 das Ribosom verlassen >> Ende der Initiation
fMet C C C Pro G G G Pro G G G fMet Pro G G G C C C Die Elongation bei der Protein-Synthese verschiedene Elongationsfaktoren (EF-Tu, EF-Ts), die GTPasen sind und GTP hydrolysieren, sind an der korrekten Anlagerung von tRNA an das Ribosom beteiligt dabei bildet sich zunächst ein Komplex zwischen der 2. tRNAPro und EF-Tu::GTP. Erst dann kann die Bindung im A-Bereich erfolgen nachdem sich der Initiationskomplex (funktionelles 70S Ribosom) gebildet hat, kann sich die zweite tRNA, die mit der entsprechenden Aminosäure beladen ist, an die A-Position anlagern. Die Auswahl der richtigen tRNA erfolgt auf Grund der richtigen Codon::Anticodon-Wechselwirkung anschließend wird GTP hydrolysiert und EF-Tu::GDP wird aus dem 70S Ribosom freigesetzt. Unter Vermittlung von EF-Ts wird EF-Tu::GTP wieder regeneriert
Die Verknüpfung der beiden Aminosäuren im Aminoacyl (A)-Bereich > Bildung der Peptidbindung Pro fMet G G G C C C Bildung der Peptid-Bindung fMet Pro G G G C C C dadurch wird im A-Bereich eine Dipeptidyl-tRNA erzeugt, während im P-Bereich eine deacylierte tRNAfMetentsteht. ursprünglich wurde angenommen, daß ein Enzym (Peptidyl-Transferase) die Peptid-Bindung im 70S Ribosom katalysiert. 1992 jedoch entdeckte man, daß die 23S rRNA diese Katalyse-Wirkung hat (keine Enzym, sondern ein Ribozym!) nach die Anlagerung der 2. Aminoacyl-tRNA an der A-Stelle, wird die neue Peptid-Bindung zwischen den bei den Aminosäuren geknüpft. Dabei greift die NH2- Gruppe der Aminosäure 2 die COOH-Gruppe der Amino- Säure 1 an der Initiator-tRNAfMet an.
das geschieht dadurch, daß das gesamte Ribosom sich exakt um die Länge eines Triplett-Codons in Richtung 3‘-Ende der mRNA bewegt (= Translokation) da die Dipeptidyl-tRNA2noch immer am 2. Codon befestigt ist, wird sie durch die Bewegung des Ribosoms vom A-Bereich in den P-Bereich verschoben, wodurch die deacylierte tRNAfMetaus dem P-Bereich ins Zytoplasma verdrängt wird fMet fMet Pro Pro G G G G G G C C C C C C GTP Gly EF-G fMet EF-G Pro A C A + GDP + Pi Translokation G G G C C C U G U Die Translokation der Dipeptidyl-tRNA2 damit der Elongationszyklus nicht stoppt und weitere Aminosäuren angeknüpft werden, muß die Dipeptidyl-tRNA2von der A-Position zur P-Position übertragen werden. das 3. Codon (UGU) der mRNA liegt jetzt im A-Bereich, das 2. Codon im P-Bereich. Diese Verschiebung benötigt ein Enzym (EF-G), das als Translokase unter GTP-Verbrauch diesen Schritt katalysiert.
NH2 Ser U G G A C C UAG = Stop-Codon Hydrolyse der Polypetidyl-tRNA- Verknüpfung NH2 Ser 50S Dissoziation der Komponenten U G G A C C 30S der Elongationszyklus schreitet solange fort, bis das Ribosom die letzte Aminosäure angefügt hat und damit das von der mRNA codierte Polypeptid fertiggestellt hat Die Termination der Polypeptid-Synthese die Terminationwird durch eines der 3 Stop-Codons (UAG - UAA - UGA) in der mRNA, für welche es keine komplementären Anticodons in der tRNA gibt, signalisiert sobald ein Stopcodon im A-Bereich erscheint, beteiligen sich 3 Terminationsfaktoren (RF = “releasing factors“) an der: 1. Hydrolyse der terminalen Peptidyl-tRNA-Bindung 2. Freisetzung des Polypeptids 3. Dissoziation des 70S Ribosoms
Die Protein-Biosynthese ist sehr energie-aufwendig > Bildung der Aminoacyl-tRNA = 2 ATP > Elongation = 1 GTP > Translokation = 1 GTP ______________________________________________ 4 ATP = 4 x energiereiche Bindungen pro 1 Peptid-Bindung = 122 kJ/mol-1 1 Peptid-Bindung hat dagegen einen Energie-Gehalt von -21 kJ/mol -1
Mehrere Ribosomen synthetisieren gleichzeitig an einer mRNA Ribosom mit kurzer Polypeptid-Kette fertig-gestellte Polypeptid-Kette Start-Codon Stop-Codon mRNA Polysomen an der mRNA
Peptidyl- Puromycin Peptidyl- Transferase Hemmung der Protein-Biosynthese durch viele Puromycin Die Wirkweise des Puromycins (aus Streptomyces alboniger) bei der Hemmung der Protein-Biosynthese Puromycin ähnelt in seiner Struktur dem 3‘-Ende einer Aminoacyl-tRNA und kann daher im A-Bereich binden Puromycin nimmt anschließend an allen Elongations-Schritten Teil bis einschließlich der Bildung der Peptid-Bindung (Peptidyl-Puromycin) Puromycin kann dagegen nicht im P-Bereich binden und dissoziiert daher als Peptidyl-Puromycinvom Ribosom ab.
die Protein-Biosynthese ist ein zentraler Vorgang in der Zelle • und daher ein Hauptangriffs-Ziel natürlich vorkommender • Antibiotica und Toxine (Tetracyclin, Chloramphenicol etc.) • wegen der Unterschiede bei der Proteinsynthese • Bakterien/Eukaryonten hemmen die meisten Antibiotica/Toxine • bei den Eukaryonten nicht!
Antibiotica and Protein-Synthese Bakterien sind für eine Reihe von mit unter letalen Infektionskrankheiten verantwortlich z. B. Tuberkulose,Pneumonia, Meningitis, Wundinfektionen, Syphilis, Gonorrhö. Vor 1940 keine effektive Behandlungsmöglichkeit mit der Entdeckung des Pencillins (hemmt bakterielle Zellwand-Synthese) änderte sich das schlagartig.Viele Antibiotica hemmen die Protein-Biosynthese. Chloramphenicol hemmt Peptidyl-Transferase > Prokaryonten Tetracyclin hemmt Initiation > Prokaryonten Streptomycin Cycloheximid hemmt Peptidyl-Transferase > Eukaryonten
A-Domäne B-Domäne Diphtherie-Toxin hemmt die Protein-Biosynthese bei Eukaryonten durch Blockierung der Translation >> Diphtherie war lange Zeit eine häufige Todesursache bei Kindern. Verursacht durch ein Toxin des Corynebakteriums diphtheriae, das sich im oberen Respirationstrakt einnistet und vermehrt >> bereits wenige mg des Gifts sind für nicht-immunisierte Personen tödlich Die A-Domäne ist ein Enzym und katalysiert die Übertragung eines ADP-Ribosyl-Restes vom NAD+ auf den Elongations- Faktor EF2 Die B-Domäne vermittelt die Aufnahme in die Zelle Hemmung der Translation Zelltod
Adäquate Häm-Menge Inadäquate Häm-Menge Regulation der Protein-Synthese inaktiv inaktiv eIF2-Phosphorylierung aktiv inaktiv Austausch von GDP zu GTP ist blockiert Translation läuft kontinuierlich ab Translation ist blockiert
Ribosomen synthetisieren auch sezernierte Proteine (z. B. Insulin)
Protein-Synthese Zytoplasma Ribosom Protein Import Gen Export Transkription mRNA (Boten-RNA) Zellkern





