Download

1 / 51
690 likes | 1.46k Views
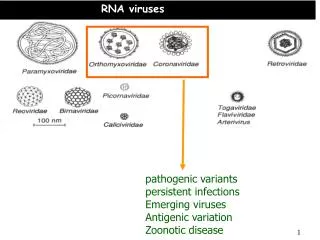
RNA viruses. Polarity (+ sense or – sense) Size of genome Segmented or not Site of replication. FAMILIES of NEGATIVE STRAND VIRUSES NON-SEGMENTED (-)STRAND VIRUSES RHABDOVIRIDAE Rabies, VSV, & Plant viruses FILOVIRIDAE - Marburg & Ebola viruses

E N D
RNA viruses • Polarity (+ sense or – sense) • Size of genome • Segmented or not • Site of replication
FAMILIES of NEGATIVE STRAND VIRUSES NON-SEGMENTED (-)STRAND VIRUSES RHABDOVIRIDAE Rabies, VSV, & Plant viruses FILOVIRIDAE - Marburg & Ebola viruses PARAMYXOVIRIDAE - Measles, Mumps, RSV, & Distemper BORNAVIRIDAE – Neurological diseases of humans and many animals SEGMENTED (-)STRAND VIRUSES ORTHOMYXOVIRIDAE - Influenza virus SEGMENTED AMBISENSE VIRUSES BUNYAVIRIDAE - Hantavirus, plant Tospovirus and Tenuivirus ARENAVIRIDAE - Lassa fever
Objectives • Segmented negative sense RNA viruses: • Orthomyxoviruses (Influenza virus A, B and C) • Bunyaviruses (include Hantavirus genus) • Arenaviruses • Double stranded RNA viruses • Reovirus (Rotavirus) • Retroviruses • HIV
What does segmented mean? • Genome in segments often representing different genes. • Segmented genomes confer evolutionary advantages. • Different strains of a virus with a segmented genome can shuffle and combine genes and produce progeny viruses or (offspring) that have unique characteristics. • This is called reassortment
Viruses with ambisense genomes • The RNA segments of some of the segmented genome viruses are ambisense, where one or more of the RNA segments each encodes two genes, one in the + sense and one in the - sense. • This is the case for the two genome segments of viruses in the family Arenaviridae and the family Bunyaviridae, such as tomato spotted wilt virus; two of the three genome segments of this virus are ambisense. • The - sense gene of an ambisense RNA is expressed by transcription of a mRNA. • The + sense gene is expressed by synthesis of an RNA complementary to the genome, followed by transcription of the mRNA for that gene.
Segmented Negative Strand RNA Viruses • Orthomyxoviridae • Three types of flu virus • Genus Influenzavirus A – 8 genome segments • Genus Influenzavirus B - 8 genome segments • Genus Influenzavirus C - 7 genome segments, no neuraminidase • Several insect-transmitted viruses • Genus Thogotovirus - 6 genome segments • Thogoto virus – tick-transmitted • Dhori virus – tick-transmitted • Batken virus – mosquito-transmitted
Segmented Negative Strand RNA Viruses • Arenaviridae • Relatively small group • Unique characteristic of encapsidating host ribosomes • Often associated with persistent infections of rodents • Several viruses associated with hemorrhagic fevers • Ambisense genomes • Bunyaviridae • Large group of 200+ viruses • Infect vertebrates, invertebrates, plants • Major component of classic “arbovirus group” • Biology usually involves vectors • Many have ambisense genomes
ORTHOMYXOVIRUSES (ORTHOMYXOVIRIDAE) • There are three groups of influenza virus: A, B and C. • Influenza A virus is most intensively studied and influenza A and B are the most important in human disease. • Influenza viruses are • pleomorphicvirions (that is, they vary in shape). • negative-sense, • single-stranded RNA • RNA genome that is SEGMENTED, eight RNA segments in influenza A. • nucleocapsid is helical • Enveloped • virions contain RNA polymerase packaged within the virus particle • Two enveloped membrane glycoproteins (figure 19): • HA - hemagglutinin - This is the attachment and fusion protein • NA - neuraminidase - This is important in release, it removes sialic acid from proteins of the virus and the host cell
Influenza etiology • Spread person-to-person by aerosol, direct or indirect contact, in water – no vectors • Incubation period 1-3 days • Causes myalgia, sore throat, fever, headache, cough which may be protracted • Symptoms typically last 2-7 days • Intensity of symptoms differs greatly depending on virus strain
Adsorption and penetration • The virus adsorbs to receptors on the cell surface and is internalized by endocytosis. • At acid pH of an endosome, HA undergoes a conformational change and fusion occurs. • Nucleocapsidsare released to cytoplasm.
Transcription, translation and replication • Nucleocapsids are transported into the nucleus. • mRNA synthesis and replication of viral RNA occurs in the nucleus. • This is very unusual for an RNA virus. Influenza virus has an unusual mechanism for acquiring a methylated, capped 5'end to its mRNAs. • A viral endonuclease (which is packaged in the influenza virus) snips off the 5'end of a host capped, methylated mRNA about 13-15 bases from the 5' end and uses this as a primer for viral mRNA synthesis. • Hence all flu mRNAs have a short stretch at the 5' end which is derived from host mRNA.
The viral RNA polymerase (transcriptase) extends the primer and copies the template into complementary plus sense mRNA and adds a poly(A) tail. • Transcription results in 8 primary transcripts, one transcript per segment. • Some segments give rise to primary transcripts which can be alternatively spliced (since influenza virus RNA synthesis occurs in the nucleus, it has access to splicing machinery), each giving rise to two alternative transcripts. • For example, the M segment gives rise to two alternative mRNAs. These code for the M1 protein and the M2 protein. Thus a single segment can code for more than one protein since the virus has access to splicing machinery. • The mRNAs are translated in the cytoplasm. • Transmembrane proteins are moved to the plasma membrane while proteins needed for RNA replication are transported to the nucleus.
Replication of RNA • RNA replication occurs in the nucleus using a virus-coded enzyme. • A full length, exact complementary copy of virion RNA is made - this + sense RNA is probably coated with nucleocapsid protein as it is made. • Full length + strand RNA is then used as a template for full-length - strand synthesis • The new - strand is probably coated with nucleocapsid protein as it is made. • New - strands can be used as templates for replication, mRNA synthesis, or packaged.
Assembly • This occurs at the plasma membrane. • Nucleocapsidsare transported out of the nucleus while envelope proteins are transported via the Golgi body to the plasma membrane. • The M1 protein interacts with both nucleocapsid and a modified region of the plasma membrane which contains the glycoproteins HA and NA. • Virus then buds out through the host cell membrane.
Reoviruses • Icosahedral symmetry • Multiple layered capsid (inner and outer capsid) • RNA is double stranded. • There are 10-12 segments (depending on the genus of the Reovirusfamily) • Due to their clinical importance in humans, we shall focus on rotaviruses.
Rotavirus replication • Rotaviruses infect cells called enterocytes at the ends of the villi (finger-like extensions) in the small intestine. • Newly synthesized (+) RNAs enter the cores, and a rigorous selection procedure ensures that each core receives one each of the 11 RNA species, a full genome complement. • Involves the recognition of a unique sequence in each genome segment. • Synthesis of (−) RNA takes place during the entry of the (+) strands into the core, VP1 again acting as the RNA polymerase. • The dsRNA of the infecting virion therefore remains intact and the mode of replication is conservative. • VP6 is added to the core, forming the second layer of the capsid. • The resulting structure is a double-layered particle similar to that derived from the infecting virion.
Transcription and translation • Double stranded RNA does not function as an mRNA - make mRNA (transcription). • The mRNAs are made by virally-coded RNA polymerase packaged in the virion. • The RNA is capped and methylated by virion packaged enzymes. • The mRNAs are translated and the resulting viral proteins assemble to form an immature capsid. • The mRNAs are packaged into the immature capsid and are then copied within the capsid to form double stranded RNAs. • More mRNAs are now made by the newly formed immature capsids.
Assembly • More proteins are made and eventually the immature capsids bud into the lumen of the endoplasmic reticulum. • They acquire a transient envelope which is lost as they mature. • This is a very odd feature of the rotaviruses. • They are released via cell lysis. • NOTE: THE ENTIRE REPLICATION CYCLE OCCURS IN THE CYTOPLASM
Proteins capsid IN = integrase matrix NC = nucleocapsid PR = protease RH = ribonucleaseH RT = reverse transcriptase SU = surface glycoprotein TM = transmembraneglycoprotein
Retrovirus Virion • Contains two copies of the RNA genome = diploid? • The two molecules are present as a dimer, formed by base pairing between complementary sequences . • The regions of interaction between the two RNA molecules have been described as a ‘kissingloop complex’. • As well as the virus RNA, the virion also contains molecules of host cell RNA that were packaged during assembly. This host RNA includes a molecule of transfer RNA (tRNA) bound to each copy of the virus RNA through base pairing. • The sequence in the virus RNA that binds a tRNA is known as the primer binding site (PBS) • Each retrovirus binds a specific tRNA.
Enzyme Activity • A number of protein species are associated with the RNA. The most abundant protein is the nucleocapsid (NC) protein, which coats the RNA, while other proteins, present in much smaller amounts, have enzyme activities. • RNA-dependent DNA polymerase (reverse transcriptase; RT) • DNA-dependent DNA polymerase • Ribonuclease H (RNase H) • Integrase • Protease
Reverse transcription takes place within the reverse transcription complex • Synthesis of both the (−) DNA and the (+) DNA begins at the 3–OH of a primer RNA. • The primer for synthesis of the (−) DNA is the tRNA bound to the genome, while the primer for synthesis of the (+) DNA is a polypurine tract (PPT) in the virus genome. • The PPT becomes accessible as a result of hydrolysis of the genome RNA from the 3 end by the RNase H, which is an enzyme that specifically digests RNA in RNA–DNA duplexes.
During synthesis of the two DNA strands, each detaches from its template and re-attaches at the other end of the template through base pairing. • The DNA that results from reverse transcription (the provirus) is longer than the RNA genome. • Each of the termini has the sequence U3–R–U5, known as a long terminal repeat (LTR), one terminus having acquired a U3 sequence and the other a U5 sequence.
A copy of the virus genome with a tRNA bound at the PBS. The reverse transcriptase begins (−) DNA synthesis at the 3 end of the tRNA. The RNase H digests the RNA from the RNA-DNA duplex. The (−) DNA attaches at the 3 end of either the same strand or the second copy of the genome. Elongation of the (−) DNA continues, while the RNase H degrades the template RNA from the 3 end as far as the PPT. LTR: long terminal repeat. PBS: primer binding site. PPT: polypurine tract (a sequence made up entirely, or almost entirely, of purine residues). R: repeat sequence. U3: unique sequence at 3 end of genome. U5: unique sequence at 5 end of genome.
5. Synthesis of (+) DNA begins. 6. The remaining RNA is degraded. 7. The (+) DNA detaches from the 5 end of the (−) DNA template and attaches at the 3 end. 8. Synthesis of both DNA strands is completed
Integration of the provirus • The provirus, still associated with some virion protein, is transported to the nucleus as a pre-integration complex . • For most retroviruses this can occur only if the cell goes into mitosis, and it is likely that mitosis-induced breakdown of the nuclear membranes is necessary for the pre-integration complex to enter the nucleus. • This means that there can be a productive infection only in dividing cells. • HIV and related viruses, however, can productively infect resting cells, as the pre-integration complexes of these viruses are able to enter intact nuclei. • One of the virus proteins still associated with the provirus is the integrase; this enzyme cuts the DNA of a cell chromosome and seals the provirus into the gap. • The integrated provirus genes may be expressed immediately, or there may be little or no expression of viral genes, in which case a latent infection has been initiated. • If a latently infected cell divides, the provirus is copied along with the cell genome and each of the daughter cells has a copy of the provirus.
Transcription and genome replication • The two LTRs of the provirus have identical sequences, but are functionally different; transcription is initiated in one and terminated in the other. • Transcription factors bind to a promoter in the upstream LTR, then the cell RNA polymerase II starts transcription at the U3–R junction. • Transcription continues into the downstream LTR. • There is a polyadenylationsignal in the R region and transcription terminates at the R–U5 junction • Each transcript is capped and polyadenylated. • Some transcripts will function as mRNA and a proportion of these become spliced; others will become the genomes of progeny virions.
Negative Strand Viruses • Contain enzymes for transcription in virion • Make mRNA prior to antigenome • Message gets capped; genome does not • Plus strand is template for minus strand genome • Makes more minus than plus strand
4. Types of Viral Genomes and Their Replication • Two events critical to viral infection: • The production of virus structural proteins and enzymes • Replication of the viral genome (dsDNA, ssDNA, dsRNA, ssRNA) Figure 3-5
dsDNA Viruses • Contain dsDNA genome • Most dsDNA viruses replicate their genomes in the nucleus of the cell • Use host’s DNA and RNA synthesizing machinery Figure 3-6 Adapted from D. R. Harper. Molecular Virology, Second Edition. BIOS Scientific Publishers, 1999.
ssDNA Viruses • Contain ssDNA genomes Adapted from D. R. Harper. Molecular Virology, Second Edition. BIOS Scientific Publishers, 1999. Figure 3-6b
ss/dsDNA Viruses (Using an RNA intermediate) • Virus carries it’s own reverse transcriptase • dsDNA enters the nucleus, forms an episome • Virus does not encode an integrase gene Figure 3-8 Adapted from D. R. Harper. Molecular Virology, Second Edition. BIOS Scientific Publishers, 1999.
RNA Viruses • Genomes may be ss or ds, (+) or (-) sense • The type of genome determines if the first step after uncoating will be translation, transcription, or RNA replication. • RNA viruses carry an RNA-dependent RNA polymerase that will synthesize viral genomes into the host cell with them. Figure 3.9: Differences between positive (+) and negative (-) sense ssRNA viral genomes.
dsRNA viruses • Contain dsRNA segmented genomes • Viral polymerase Figure 3.10: List of dsRNA viruses and their replication strategy. Adapted from D. R. Harper. Molecular Virology, Second Edition. BIOS Scientific Publishers, 1999.
+ssRNA Viruses • Contain +ssRNA nonsegmented genomes • The RNA in the virus particle functions as mRNA • Viral mRNA is recognized by cellular translational machinery • Contain a viral RNA-dependent RNA polymerase in order to replicate viral genomes Figure 3.11: List of +ssRNA viruses and their replication strategy. Adapted from D. R. Harper. Molecular Virology, Second Edition. BIOS Scientific Publishers, 1999.
-ssRNA viruses • Contain -ssRNA segmented or nonsegmented genomes • Contain a viral RNA-dependent RNA polymerase gene Figure 3-12b Figure 3-12a Adapted from D. R. Harper. Molecular Virology, Second Edition. BIOS Scientific Publishers, 1999.
Viruses with ssRNA Genomes That Use a dsDNA Intermediate to Replicate • Unique biology • Viral genome is reverse transcribed and integrated as a cDNA into the host’s chromosome Adapted from D. R. Harper. Molecular Virology, Second Edition. BIOS Scientific Publishers, 1999. Figure 3-13