Download
1 / 55
550 likes | 644 Views
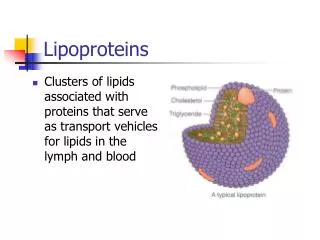
Lipoproteins 脂蛋白. Fat Metabolism during Exercise. Adipose Tissue - Triacylglycerol Fatty Acid + Glycerol circulation Re-esterification 再酯化 : FA not released into circulation, but used to form new TG within adipose tissue Intramuscular Triacylglycerol (IMTG)

E N D
Ex Nutr c7-fat Lipoproteins 脂蛋白
Ex Nutr c7-fat Fat Metabolism during Exercise • Adipose Tissue - Triacylglycerol • Fatty Acid + Glycerol circulation • Re-esterification 再酯化: FA not released into circulation, but used to form new TG within adipose tissue • Intramuscular Triacylglycerol (IMTG) • Hormone Sensitive Lipase (HSL) • Circulation Triacylglycerol • Via Lipoprotein Lipase
Ex Nutr c7-fat Fig 6.1 overview of fat metabolism
Ex Nutr c7-fat What Limits Fat Oxidation • Lipolysis in Adipocytes • Removal of FAs from fat cell • Transport of fat by the bloodstream • Transport of FAs into the muscle cell • Transport of FAs into the Mitochondria • Oxidation of FAs in the β-oxidation pathway and TCA cycle
Ex Nutr c7-fat Lipolysis in Adipocytes • HSL – inactive form and active form • Sympathetic nervous system • Epinephrine • Insulin • Fate of glycerol • bloodstream – Liver for gluconeogenesis • Not used in adipocytes, because glycerolkinase very low (glycerol glycerolphosphate glycolysis)
Ex Nutr c7-fat Lipolysis in adipocytes • Fate of fatty acid • Re-esterification: 70% at rest • FA-binding proteins (FABP), transport FA to cell membrane • Exercise:↓re-esterification, ↑lipolysis ↑ FA in blood • Lipolysis in excess of demand for FA at rest and during exercise • Glycerol-P for re-esterification in adipocytes from glycolysis pathway
Ex Nutr c7-fat Removal of FAs from fat cell and Transport of fat by the bloodstream • Blood flow to adipose tissue • Albumin concentration and available binding sites • A transporter of FAs • 3 site for FAs • Typical plasma concentration 0.7 mM (45 g/L) • Albumin bind to albumin-binding proteins in target tissue, facilitate FA release and uptake • Maximal FA concentration in blood – 2 mM • Free FA toxic • Protective mechanism: incorporated into VLDL in liver • Lipoprotein TG <3% energy during prolonged ex • LPL activity ↑after training, ↑fat oxidation
Ex Nutr c7-fat Transport of FAs into the muscle cell • Plasma membrane FA binding protein (FABPpm) • FA transporter – FAT/CD36 • Fatty acid translocase • Saturated at 1.5 mmol/l - FA • Muscle contraction – translocation FAT/CD36 • Cytoplasmic FA binding protein (FABPc)
Ex Nutr c7-fat Fig 7.4 transport of glucose and FA
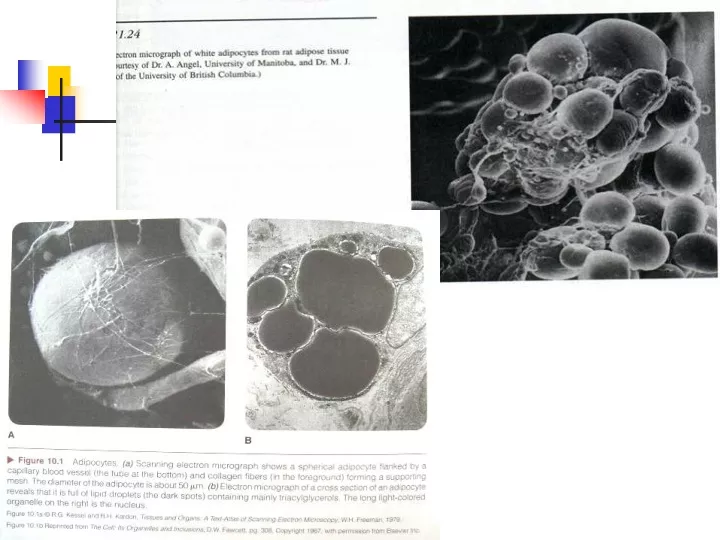
Ex Nutr c7-fat Intramuscular Triacylglycerol • Lipid droplets, usually located adjacent to mitochondria • Important energy source during exercise • Located closer to Mitochondria in trained muscle • HSL • FA released bound to FABPc until transported into mitochondria
Ex Nutr c7-fat Fig 7.5 Electromyograph of skeletal muscle. Lipid droplet (li); mitochondria (mi); glycogen (gl)
Ex Nutr c7-fat Transport of FAs into Mitochondria • Acyl-CoA Synthetase • CPT I & CPT II Carnitine Palmitoyl Transferase I & II • Short- and Medium-Chain FAs diffuse freely into mitochondrial matrix • Fatty acyl-CoA in mitochondrial matrix is subjected to beta-oxidation • Acetyl-CoA for TCA cycle
Ex Nutr c7-fat Fig 7.6 Transport of FA into mitochondria
Ex Nutr c7-fat Fig 7.7 beta-oxidation
Ex Nutr c7-fat Fat As A Fuel During Exercise • At Rest • Lipolysis regulated by hormones • FA concentration 0.2-0.4 mM in blood • During moderate exercise • ↑Lipolysis ~3X, ↑ blood to adipose tissue 2X, ↓re-esterification 0.5X, ↑ blood flow to muscle • FA in blood decrease in the first 15 min: rate of uptake by muscle > rate of appearance by lipolysis • FA increase thereafter, may reach 1 mM • Higher intensity exercise • Rise in plasma FA very small or absent • Lactate ↑re-esterification in adipocyte, ↓FA release
Ex Nutr c7-fat Lactate ↑re-esterification in adipocyte,↓FA release
Ex Nutr c7-fat Fat Oxidation,Exercise Duration and intensity • ↑ Fat oxidation when exercise duration ↑ • Fat Oxidation rate at 1.0 g/min after 6 hr running • Fat oxidation rate can reach 1.5 g/min after high-fat meal • Low to moderate intensity – Fat dominated • >75%VO2max – Fat oxidation rate inhibited • Maximal fat oxidation rate usually 62-63% VO2max
Ex Nutr c7-fat Fig 7.8 Fat oxidation as function of exercise intensity
Ex Nutr c7-fat Fat Oxidation and Exercise Intensity • Infusion of TG (Intralipid) and heparin 肝素during higher intensity exercise ↑FA oxidation rate • But still lower than at moderate intensity • Even plasma FA concentration same as that at moderate intensity • During higher intensity exercise, oxidation of long-chain FA impaired, but oxidation of medium-chain FA unaffected • ↑glycolytic flux, ↑pyruvate and acetyl-CoA, inhibit CPT-1 • Carnitine-dependent FA transport may be limiting factor for FA oxidation during exercise • Medium-chain FA do not need transport system across mitochondria
Ex Nutr c7-fat Fig 7.9 substrate utilization at different exercise intensities
Ex Nutr c7-fat Fig 7.10 Lipid-heparin infusion significantly ↑plasma [FA] and fat oxidation rate at 85% VO2max. The fat oxidation rate with infusion is still lower than that in 65% VO2max (not shown), Fat infusion ↓CHO oxidation rate by ↓ muscle glycogen utilization (not shown)
Ex Nutr c7-fat Fat Oxidation and Aerobic Capacity • Endurance training↑fat oxidation contribute to total energy expenditure • ↑ Mitochondria density, ↑ Oxidative enzyme • ↑ Capillary density ↑FAs, oxygen to muscle • ↑ FABP • ↑ CPT • After training, rate of lipolysis at same absolute exercise intensity NOT affected • After training, rate of lipolysis at same relative exercise intensity NOT affected • ↑IMTG lipolysis contribute to ↑whole-body lipolysis
Ex Nutr c7-fat Fig 7.11 Whole-body lipolysis in trained and untrained subjects
Ex Nutr c7-fat Fat Oxidation and Diet • High CHO, low fat diet : ↓fat oxidation • High fat, low CHO diet: ↑fat oxidation • Fat oxidation still higher after 5 days of high-fat diet and then 1- day high-CHO diet • Compared to 6-day high-CHO diet • 1-day high-CHO diet replenish muscle glycogen, similar levels in both groups • Not due to substrate availability • Adaptations at muscular level may occur within 5 days • Chronic diet may affect metabolism
Ex Nutr c7-fat Response to Carbohydrate Feeding • Ingestion of CHO before exercise, ↑ Insulin secretion • ↑CHO oxidation, ↓Fat oxidation • FA availability only partially responsible • Glucose feeding 1 hr before exercise ↓plasma FA concentration and ↓ Fat oxidation • Infusion of Intralipid and heparin only partially restored fat oxidation • Study using labeled Long- and Medium-chain FA indicated carnitine-dependent FA transport into mitochondria also responsible
Ex Nutr c7-fat Regulation of CHO & Fat Metabolism • ↓Muscle glycogen breakdown with ↑plasma FA concentration • Infusion of intralipid • Fat feeding in combination with heparin infusion (↑lipoprotein lipase and hepatic lipase activities) • Problems in control group • FA concentration very low, <0.2 mM • ↑glycogen breakdown in control group, rather than observed glycogen-sparing effect in high-fat group
Ex Nutr c7-fat Regulation of CHO & Fat Metabolism –glucose- fatty acid cycle • ↑plasma FA, ↑FA uptake, ↑acetyl-CoA and citrate • ↑acetyl-CoA/CoA inhibit pyruvate dehydrogenase • ↓pyruvate acetyl-CoA • ↑citrate inhibit phosphofructokinase • After citrate diffuse into sarcoplasma • ↓glycolysis accumulation of glucose-6-P in sarcoplasma ↓hexokinase, ↓muscle glucose uptake • ↑FA availability ↑NADH ↓intramuscular Pi and AMP • Pi and AMP stimulate glycogen phosphorylase • ↑FA availability ↓muscle glycogen breakdown
Ex Nutr c7-fat Fig 6.14 Glucose-FA cycle, or Randle cycle
Ex Nutr c7-fat Regulation of CHO & Fat Metabolism –recent theory • Fat dose NOT regulate CHO metabolism • It is CHO that regulates fat metabolism • ↑glycolysis ↓fat oxidation • ↑acetyl-CoA in muscle during high-intensity ex • Some acetyl-CoA malonyl-CoA by acetyl-CoA carboxylase • CPT1 inhibition ↓FA transport to mitochondria • By ↑ Malonyl-CoA • Also by ↓ intramuscular pH during high-intensity ex • ↑acetyl-CoA bind to carnitine ↓free carnitine, ↓cartinine for FA transportation • Fat oxidation mainly influenced by fat availability and rate of CHO oxidation
Ex Nutr c7-fat Fig 6.15 Glucose FA cycle reversed
Ex Nutr c7-fat Fat Supplementation and Exercise • Ingestion of Long-Chain Triacylglyerols • Dietary LCFA TG in chylomicron • Breakdown of TG in chylomicron very low by muscle during exercise, should avoid fat intake • Replenishment of IMTG stores AFTER exercise • Some sports bars or energy bars contain high fat • Ingestion of Medium-Chain Triacylglyerols • C8 or C10, very low in natural foods, mainly used as supplements • Most studies showed no effect on performance • Large ingestion (> 15 g/h) may cause GI problems • Ingestion of Fish Oil • Omega-3 FAs • More omega-3 FAs incorporated into membrane, may improve membrane characteristics and function
Ex Nutr c7-fat Effects of Diet on Fat Metabolism and Exercise Performance • Fasting • ↑lipolysis, ↑plasma FA • ↓ endurance performance, even with CHO ingestion during exercise • Effects of Short-term high-fat diet • ↑lipolysis, ↑plasma FA ,↑Fat oxidation rate, ↑ Ketone bodies 酮體(5X after 5 days of high-fat diet) • ↓ endurance performance, ↓ muscle glycogen
Ex Nutr c7-fat Effects of long term high-fat diet (several weeks) on performance • 60-65% VO2max, may be impractical • Maintain time to exhaustion at 60-65% VO2max, despite muscle glycogen ↓by 50% • CPT↑ 44%, hexokinase ↓46% • Relatively low intensity, reduced CHO stores may not be limiting factor • 7 wk high-fat + 1 wk high-CHO vs 8 wk high-CHO • Time to exhaustion at 81% VO2max↑from wk7 to wk 8 • Still less than 8-wk high-CHO group • Suboptimal adaptation to training • High-fat diet • may only helpful in low-intensity exercise • Negative effect on general health • Not recommended for athletes
Ex Nutr c7-fat Fig 7.17 high-fat diets and performance during training in humans
Ex Nutr c7-fat Fat adaptation impair glycogenolysis • Most fat adaptation studies (days or weeks of high-fat diet) show no effect on endurance performance • Despite ↑fat oxidation, ↓CHO oxidation • High-fat diet ↑pyruvate dehydrogenase kinase, ↓pyruvate dehydrogenase • At rest, throughout 70% VO2max exercise, 1-min sprint • ‘glycogen sparing’ effect of high-fat diet actually impairment of glycogenolysis
Ex Nutr c7-fat Spriet LL, JSS 2004