Download

1 / 36
590 likes | 1.59k Views
Immunoglobulin Gene Organization and Expression. W. Robert Fleischmann, Ph.D. Department of Urologic Surgery University of Minnesota Medical School rfleisch@umn.edu (612) 626-5034. Objectives. Understand how the immunoglobulin genes are organized in the DNA.

E N D
Immunoglobulin Gene Organization and Expression W. Robert Fleischmann, Ph.D. Department of Urologic Surgery University of Minnesota Medical School rfleisch@umn.edu (612) 626-5034
Objectives • Understand how the immunoglobulin genes are organized in the DNA. • Understand the basis for diversity of immunoglobulins. • Understand the mechanisms by which the variable regions undergo gene rearrangement. • Understand the mechanism of class switching. • Understand how immunoglobulin genes are expressed. • Understand the medical consequences of the activity of genetic rearrangement mechanisms.
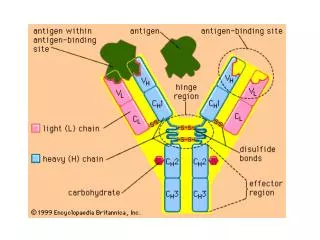
Light chain (kappa or lambda) Heavy chain (isotypic region: , , , or) General Structure of an Antibody Light chains: Kappa chains are found twice as frequently in Abs as are lambda chains. Heavy chains: These undergo class switching as the B cell undergoes differentiation.
The Conundrum • The human genome contains an estimated 20,000 to 25,000 genes that encode mRNAs for proteins. • To protect us, our immune system has the ability to produce about 1010-1011 different antibodies. • If every gene encoded a different antibody, the genes would encode only 1 millionth of the antibodies needed. • How can our immune system produce so many different antibodies with so few genes?
Features of the Antibody Genes • Antibodies are composed of heavy and light chains. • As is seen for most eukaryotic genes, the heavy and light chains of the immunoglobulin genes are each composed of segments (exons) that must be joined together to form the immunoglobulin genes. • For immunoglobulin genes, the joining of a number of the exons occurs via a rearrangement of the gene segments at the level of the DNA, rather than at the level of the mRNA. • There are multiple copies of each of the various segments of the heavy and light chains of the immunoglobulin genes, with one of each of these segments becoming sequentially rearranged to form the heavy and light chain genes.
The Keys to Antibody Diversity • Antibody diversity is generated during genetic rearrangement by mixing and matching one of each of the various gene segments for the heavy and light chains in a combinatorial manner. • Antibody diversity is generated by errors incorporated at the joining sites for the various segments of the heavy and light chains. • Antibody diversity is generated by hypermutation in one of the gene segments (variable regions) of the heavy and light chains during proliferation of B cells. • Antibody diversity is generated by mixing and matching heavy and light chains in a combinatorial manner.
Overview of Immunoglobulin Expression at Various Stages of B Cell Maturation
The Immunoglobulin Heavy Chains • There are two identical immunoglobulin heavy chains in each antibody. • Each of the immunoglobulin heavy chain genes is assembled from V, D, J, and C gene segments. • There are multiple C gene segments (constant regions) that give rise to different isotypes. • IgD • IgM • IgG: 4 heavy chains, IgG1, IgG2, IgG3, IgG4 • IgE • IgA: 2 heavy chains IgA1, IgA2
Heavy Chain Diversity • 39 V gene segments • 23 D gene segments • 6 J gene segments 5,382 combinations of heavy chain VDJ segments
The Immunoglobulin Light Chains • There are two types of immunoglobulin light chains. • light chain • light chain • Each of the immunoglobulin light chain genes is assembled by the rearrangement of V, J, and C gene segments.
Light Chain Diversity • light chain diversity • 40 V gene segments • 5 J gene segments • 1 C gene segment • light chain diversity • 30 V gene segments • 4 J gene segments • 4 C gene segments 200 combinations of light chain VJC segments 480 combinations of light chain VJC segments 680 combinations of light chain VJC segments
Antibody Diversity from Rearrangements • From heavy chains: 5,382 combinations • From and light chains: 680 combinations 3.6 x 106 Combinations of light and heavy chains This is far short of the 1010-1011 different Antibodies that are postulated to occur.
How Does Rearrangement Occur? • Rearrangement occurs between specific sites on the DNA called recombination signal sequences (RSSs). • Rearrangement is catalyzed by two recombination-activating genes: RAG-1 and RAG-2.
Recombination Signal Sequences • The specific recognition sequences, called recombination signal sequences (RSSs), indicate the sites of recombination. • These signal sequences composed of a 7 bp sequence and a 9 bp sequence are separated by one turn of the DNA (12 bp spacer) or by two turns of the DNA (23 bp spacer). • The RSSs are inverted repeats that allow the DNA to form a stem-loop with the RSSs aligning on the stem.
Rearrangement Process Site of P and N nucleotide additions
P-Nucleotide AndN-Nucleotide Additions • Cleavage of the RSSs by RAG leaves single-stranded regions. • These single-stranded regions are copied to form a hairpin loop by addition of P-nucleotides (P for palindromic nucleotides). • Additional N nucleotides (N for any nucleotide) may be added. • The sequences are joined.
Consequences of Rearrangement and P- and N-Nucleotide Addition • Positive consequence of imprecise joining of Ig gene segments • A productive rearrangement occurs if the number of nucleotides added across the joining region allows the genetic code to be read in phase. • This results in the generation of additional diversity. • Negative consequence of imprecise joining of Ig gene segments • A nonproductive rearrangement occurs if the number of nucleotides added across the joining region causes the genetic code to be read out of phase (the majority of times). • This results in an incomplete antibody (run into stop codons) • The B cell may be able to productively rearrange the immunoglobulin gene on the other chromosome. • Otherwise, it will result in the death of the B cell.
Alleleic Exclusion • Alleleic exclusion occurs when only one of two alleles is expressed. • This is the case with immunoglobulin molecules. • This ensures that a given B cell will make antibody molecules with only a single specificity. • Antibodies may be made from maternal and paternal chromosomes:HM:LM; HP:LM; HM:LP; HP:LP. Note: If the first allele makes a non-functional antibody, the second allele will undergo rearrangement.
Immunoglobulin Molecules Expressed on Mature B Cells • Mature (but not activated) B cells initially express IgD and IgM on their external cell membranes. • The choice of IgD versus IgM occurs at the level of processing of mRNA, so a given B cell can both express IgD and IgM. • As mature B cells are activated to divide and differentiate by their cognate antigen, they switch from membrane-bound IgD and IgM to secretory IgM. • This switch occurs at the level of processing of mRNA transcripts. • As they continue to divide and differentiate, they may undergo additional class switching: IgM => IgG => IgE => IgA. • These switches occur at the level of rearrangements of the DNA.
Expression of Membrane-Bound IgD and IgM • An initial pre-mRNA transcript is produced. • Importantly, the pre-mRNA transcript has two poly-A sites. • If the second polyadenylation site is read, then the mRNA for membrane-bound IgM is generated by splicing. • If the fourth polyadenylation site is read, then the mRNA for membrane-bound IgD is generated by alternate splicing.
Structures of Membrane IgM and Secreted IgM Note the many hydrophobic amino acids in the trans- membrane sequence. Note the cysteine for creation of S-S bonds between IgM antibodies.
Expression of Membrane-Bound IgM and Secreted IgM • The initial pre-mRNA transcript is synthesized. • Importantly, the pre-mRNA transcripts have two poly-A sites within the Cµ gene segment. • If the M1, M2 exons are spliced out, the mRNA for secreted IgM is produced.
Class Switching With Activated B Cell Differentiation And Division • After activation, B cells switch from membrane-bound IgM and IgD to secreted IgM by differential splicing. • As the activated B cells continue to differentiate and divide, they class switch to production of IgG by DNA rearrangement. • Activated B cells may continue to class switch to production of IgE or IgA by DNA rearrangement.
Mechanism of Class Switching • With activation of the B cell, class switching can occur. • At the level of the DNA, a looping event occurs that cuts out the constant regions for IgM and IgD. • This leads to the production of IgG mRNAs. • If further looping out occurs, the mRNAs for IgE or IgA are produced.
Cytokine Effects on Class Switching Certain cytokines affect class switching: IFN- => IgG2a IL-4 => IgG1, IgE IL-5 => IgE
Antibody Diversity by Hypermutation • After exposure to its cognate antigen, the mature B cell is activated to proliferate. • As the B cell proliferates, mutations accumulate in the immunoglobulin gene by a process called somatic hypermutation. • These mutations are concentrated in the variable region. • The mutations give greater antibody diversity. • Some of the mutations will lead to an antibody that binds the antigen more firmly (affinity maturation). • If the mutations lead to less Ab affinity, the B cell dies.
Review of Immunoglobulin Expression at Various Stages of B Cell Maturation
Diseases Associated With Expression of Immunoglobulin Genes • Burkitt’s Lymphoma • Acute Lymphocytic Leukemia • Acute Myelogenous Leukemia • Chronic Lymphocytic Leukemia • Chronic Myelogenous Leukemia
Diseases Involving Translocations of Oncogenes Onto Immunoglobulin Genes • Chronic Myelogenous Leukemia: • Philadelphia chromosome (novel chromosome 22) • This results from a reciprocal translocation of chromosomes 9 and 22. • This places a proto-oncogene under the control of the light chain gene promoter on chromosome 22. • Burkitt’s Lymphoma: • A reciprocal translocation of chromosomes 8 and 14. • This places c-myc under the control of the heavy chain promoter on chromosome 14.
Medical Uses of Antibodies • Monoclonal antibodies are used for treatment of certain diseases.
Anti-Idiotopic Antibody • The concept is to make an antibody that mimics an antigen. • This is not in use yet. • It could be used to generate antibodies against antigens. • Antigens that are not readily recognized: a polysaccharide molecule • Antigens that are not readily recognized and toxic: the lipid A region of bacterial endotoxin. • The anti-idiotype antigen could then be used as a vaccine.
Generation of Monoclonal Antibodies • B cells from recently immunized mice are harvested from spleens and fused with mouse myeloma cells. • The myeloma cells have been cloned to resist a specific toxic drug. • Only myeloma cells and fused myeloma cells survive treatment with the toxin. • The surviving cells are cloned and tested for production of antibody to the antigen used to immunize the mice. • The identified clone is cultured and harvested for monoclonal antibody.
Production of Humanized Monoclonal Antibodies • Harvest mouse embryonic stem cells and KO the genes for the mouse heavy and light chains. • Transfect the KO mouse cell with a human artificial chromosome that encodes the human heavy and light chains. • Inject into a mouse blastocyst to generate a chimeric mouse with 1 normal mouse and one KO mouse chromosome for antibody genes. • Interbreed progeny mice to obtain a mouse that is homozygous for the KO mouse chromosome and the human artificial chromosome. • Inoculate mice with antigen as for generating monoclonal antibody. Immunize with antigen. Harvest spleen cells. Fuse with myeloma cells.