Download

1 / 32
340 likes | 451 Views
Structures of Mismatch Replication Errors Observed in a DNA Polymerase. Sean J. Johnson and Lorena S. Beese Biochemistry 4000 Tracy Wang. High-fidelity DNA polymerases maintain replication accuracy through:.

E N D
Structures of Mismatch Replication Errors Observed in a DNA Polymerase Sean J. Johnson and Lorena S. Beese Biochemistry 4000 Tracy Wang
High-fidelity DNA polymerases maintain replication accuracy through: • Select for correct base pairing, while strongly discriminating against mismatched bases ( prior to covalent incorporation) • Stalling of the polymerases, thereby favoring subsequent mismatch excision (after incorporation)
Klenow Fragment • DNA polymerase I • Highly processive • Fingers, Thumb and Palm
Taq polymerase • Closed conformation • dNTP bound • Open conformation • No incoming nt.
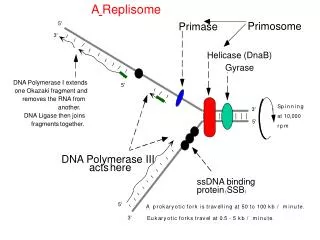
Model system • Use the thermophilic Bacillus DNA polymerase I fragment ( BF). • BF is a high-fidelity Family A polymerase. • Structural homology to the Klenow Fragement of E.coli (KF) and T. aquaticus (Taq) polymerases.
Model system • Insertion site (n position ) • Catalytic site • Preinsertion site • Postinsertion site (n-1 position ) • DNA duplex binding region (n-2,…, n-5 positions )
Replication mechanism • Transfer of the n template base from the pre-insertion site to the insertion site: - open to closed conformation - hinge-bending motion • Base pairs with an incoming base, covalent incorporation takes place. • Newly synthesized base moves into the post-insertion site. • The DNA in the duplex binding region translocates by one base pair. • The next template base moves into the pre-insertion site.
Cognate G•C base pair bound at the postinsertion(n-1) site. • Hydrogen bond between Asp830 and the 3’ primer terminus. • Arg615 and Gln797 interact with hydrogen bond acceptor atom located in the DNA minor groove.
Objective • To obtain and solve the structure of all 12 possible mismatches captured at the active site of a DNA polymerase. • To understand the mechanisms that lead to mismatch-induced stalling of the polymerase.
Crystallographic capture of mismatches • Crystals with mismatches were obtained either by enzymatic incorporation in the crystals (E), or by cocrystallization of DNA duplexes that contain a mismatch at the primer terminus (C). • In the presence of Mg2+, accurate DNA replication is obtained. • In the presence of Mn2+, enzymatic incorporation of mismatches is allowed. • E: use mutagenic reaction conditions (MgSO4 MnSO4 ) • C: BF cocrystallized with DNA substrates
Results • 12 possible covalently incorporated DNA mismatches. • Six are placed at the post-insertion site and are well ordered (G•T, T•G, T•T, C•T, A•G, G•G). • Three are placed at the post-insertion site but are too disordered for interpretation (T•C, A•C, C•A). • Three are frayed (A•A, C•C, G•A). Primer strand is at the insertion site Template strand is at the pre-insertion site
Results • Structure of each mismatch is distinct. • Some structures had been previously observed, others do not. • The type and degree of disruptions vary depending on the identity of the mismatch.
Mismatch-induced disruptions at the active site • Displacement of the template strand • Repositioning of Tyr714 • Block the template pre-insertion site due to the rearrangement • B form DNA at the active site rather than A form • The catalytic site is undisturbed
G adopts a conformation similar to that of a cognate base intact catalytic site • Wobble conformation, T is positioned toward DNA major groove • Minor groove interaction with Gln797 is lost displacement of template strand • Primer base G rotates 180 into a syn conformation, template base remains in an anti conformation helical width is closer to a Watson-Crick base pair • More closely resembles the G•T complex • Template strand is displaced • N3 is moved from the minor groove interaction between Arg615 and N3 is lost
T•C, A•C : disordered mismatches • The disorder is confined primarily to the template strand • Disorder on the primer strand is localized to the primer base • A mixture of A and B forms of DNA can be discerned in the DBR. • Protein adopts a distorted open conformation in which the pre-insertion site is blocked.
The primer terminus is displaced • Interaction between Asp830 and the primer 3’ hydroxyl at the catalytic site is disrupted • The DNA backbone of the template strand is undisturbed
wobble conformation. • The primer base lifts up into the DNA major groove, and the template base rotates slightly toward the minor groove. • Shift of the 3’ hydroxyl breaking the hydrogen bond with Asp830 disrupting assembly of the catalytic site. • Not a wobble conformation. • Base pair directly opposite each other • Significant opening in the minor groove • Bridging water molecule
Displacement of the template strand. • Blocking of the pre-insertion site. • Displacement of the primer strand. • Disruption of the catalytic site.
Both bases maintain an anti conformation increase of the helical width extensive movement of both template and primer strands. • Altering the interaction with Asp830, but without breaking it. • Wobble conformation. • An additional water-mediated H-bond. • Template is displaced • Interaction between Arg615 and the primer base is lost • Sugar ring of T is disordered primer terminus is displaced, but interaction between Asp830 and primer 3’ hydroxyl may be retained to some degree.
Structure is frayed. • The primer base is bound at the insertion site. • The template base is bound at the pre-insertion site.
A•A, C•C, G•A: frayed mismatches. • Do not bind in the post-insertion site. • Tyr714 blocks access of the template base to the insertion site. • Tyr714 also prevents the primer base from stacking to the DNA helix. Primer base stacks against Phe710. • Catalytic site is completely disrupted.
Mismatch extensions • By transferring BF.DNA cocrystals containing a mismatch at the 3’ primer terminus into reaction buffers containing nucleotides complementary to the template strand. • By soaking in different combinations of nucleotides in the presence of Mg2+ to control the final location of the mismatch
Results • The A•G, T•T, T•G, and C•C mismatches failed to extend. • The G•T, C•T, and G•G mismatches were all successfully extended. • For the T•C mismatch, DNA is too disordered to permit interpretation.
G•T Extension • At n-2 position: wobble • At n-2 and n-3: disrupt A to B form transition, similar to the disruptions observed when G•T is at n-1 disrupted active site • At n-4: partially restores the normal DNA structure mixture of both a disrupted and an undisrupted active site • At n-3 and n-4 : the wobble inverts inconsistent with the interbase H-bonding geometry associated with the major tautomeric form of nt. tautomeric shift or ionization of the mismatch • At n-6: fully restores the DNA conformation
C•T Extension • Extended in a single round of replication • At n-2 position: fully restores the catalytic site.
Summary • The high-resolution structures of all 12 possible covalently incorporated DNA mismatches at the active site of the BF polymerase were obtained. • The mismatch-induced disruptions can be divided into four broad categories. • Disruption of template strand and pre-insertion site. • Disruption of primer strand and assembly of catalytic site. • Disruption of template and primer strands. • Fraying of DNA at insertion site. • The polymerase retains a short-term memory of the mismatch incorporation event.
Discussion • Category of mismatch-induced disruptions differences in mismatch extension efficiencies ? • G•T: disruption may account for the reduced extension efficiency ten-fold to a thousand-fold. G•G: more than 100-fold slower than that of G•T. • C•T: can be extended readily due to the flexibility of DNA. • A•G: efficiencies are reduced by up to million-fold.
Discussion • A heteroduplex can adjust its size in order to translocating through the DBR. • The structural adaptations are confined primarily to the DNA, with some local protein side chain motions. • Exonucleolytic excision of mismatches requires dissociation of the heteroduplex from the polymerase: A number of the structures show diminished interactions. • Some mismatch conformations have not been previously described in a DNA heteroduplex. - Structural adaptations are not confined to simple conformational rearrangements. • Each equivalent mismatch interacts in a unique manner with the protein. (e.g. G•T, T•G)
Future work • Basis for future biochemical and structural studies. • Study the structural changes of a given mismatch under different conditions (pH, sequence context). • To see if these mechanisms can be applied to other DNA polymerases and even RNA polymerases.
References • Johnson, S. J. and Beese, L. B. Cell 2004, 116, 803-816.