Download
1 / 189
1.89k likes | 2.11k Views
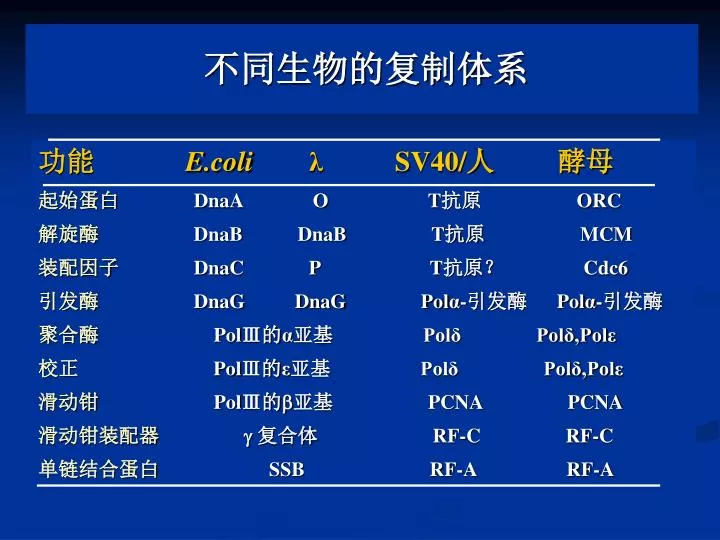
不同生物的复制体系. 功能 E.coli λ SV40/ 人 酵母 起始蛋白 DnaA O T 抗原 ORC 解旋酶 DnaB DnaB T 抗原 MCM

E N D
不同生物的复制体系 功能 E.coliλ SV40/人 酵母 起始蛋白 DnaA O T抗原 ORC 解旋酶 DnaB DnaB T抗原 MCM 装配因子 DnaC P T抗原? Cdc6 引发酶 DnaG DnaG Polα-引发酶 Polα-引发酶 聚合酶 PolⅢ的α亚基 Polδ Polδ,Polε 校正 PolⅢ的ε亚基 Polδ Polδ,Polε 滑动钳 PolⅢ的亚基 PCNA PCNA 滑动钳装配器 复合体 RF-C RF-C 单链结合蛋白 SSB RF-A RF-A
原核生物和真核生物DNA复制的比较 相同点: 1)半保留复制方式 2)半不连续复制 3)DNA 螺旋酶, SSBP 4)RNA 引物 不同点: 1) 复制起点(单、多) 2)复制子(大小、多少) 3)复制叉移动的速度 4)冈崎片段的大小 5)端粒和端粒酶 6)DNA聚合酶
复制子的大小 : 酵母 平均 40 kb 脯乳动物 平均 100 kb 原核生物 >1000 kb 冈崎片段 原核生物 1000-2000 nt 真核生物 100-200 nt 复制速度 原核生物50,000bp/min (750/sec) 真核生物 3,000bp/min (50/sec) DNA聚合酶 真核生物 五种,α、β、γ、δ和ε 原核生物 三种 ,polⅠ、Ⅱ、Ⅲ 引物酶作用特点 真核细胞 DNA引物酶和polα紧密偶联, 原核细胞 引发酶和解旋酶偶联,形成复制体的一部分。
RNA合成的基本特征: • 5’→3’方向; • 底物三磷酸核苷酸(NTP) • 不对称转录,以单链DNA为模板。 • 不需要引物,合成是连续的。 • 对一个基因组来说,转录只发生在一部分基因,且每个基因的转录都受到相对独立的控制。
典型原核生物启动子的结构 原核生物启动子的结构特点 -35 -10 转录起点 TTGACA 16-19bp TATAAT 5-9bp
I 型: II 型 III型 -170 -160 -150 -140 -130 -120 -110………..-60 -50 -40 -30 -20 -10 +1 +10 +20 上游控制区 核心启动子 真核生物 RNA pol 启动子
原核生物转录初始的tRNA和rRNA都不是最初的转录产物 因为: (1) 5’端都是单磷酸,而原始的转录产物5’三磷酸; (2) 分子比初始转录物小; (3) tRNA含有特殊的碱基,这些碱基只有通过一些化学修饰才可以得到。
真核mRNA的加工和成熟 5’加帽 3’加尾 特殊碱基的修饰 内含子的切除
转录(transcription):以DNA为模板合成RNA的过程。转录(transcription):以DNA为模板合成RNA的过程。 • 转录所需的酶叫做依赖DNA的RNA聚合酶。 • 转录产生初级转录物为RNA前体(RNA precursor),必须经过加工过程变为成熟的RNA。
模板链, 无意义链:在DNA的两条多苷酸链中只有其中一条链作为模板。 • 编码链,有意义链: DNA双链中另一条不做为模板的链; • 不对称转录:RNA的转录合成是以DNA的一条链为模板而进行的。
第一节 转录酶 • 第二节 启动子 • 第三节 终止子 • 第四节 转录的机制 • 第五节 转录产物的后加工 • 第六节 RNA的剪接
第一节 转录酶1 原核生物的RNA聚合酶2 真核生物的RNA聚合酶
RNA合成的基本特征: • 5’→3’方向; • 底物三磷酸核苷酸(NTP) • 不对称转录,以单链DNA为模板。 • 不需要引物,合成是连续的。 • 对一个基因组来说,转录只发生在一部分基因,且每个基因的转录都受到相对独立的控制。
1 原核生物的RNA聚合酶(RNA polynerase) 1.1 RNA pol与DNA pol作用的相同点: DNA模板,Mg2+,4种三磷酸核苷。 • 不同点: (1)RNA pol没有任何校对功能; (2)能起始新的RNA链。
1.2组成 • 大肠杆菌RNA聚合酶由多亚基组成, α2ββ’ωσ称为全酶, 480 KDa • 亚基与全酶结合疏松,很容易与全酶分离。
1.3 各亚基的功能 β亚基:由rpoB编码,结合底物(NTP及新生RNA链),进行聚合作用。 β′亚基:由rpoC编码,可能与模板结合。 α亚基:由rpoA编码,可能参与全酶的组装及全酶识别启动子,还参与RNA聚合酶与一些调控因子间的作用。 ω 亚基:约10KD,作为核心酶,具体功能不清楚。 σ亚基/因子:使全酶识别启动子并与之结合,也看作一种辅助因子。
1.4 σ亚基/因子 • σ因子在RNA聚合酶识别启动子的过程中起关键作用,σ因子可重复使用。 • 不同的因子可识别不同的启动子
因子更替的现象 • 在细菌受到外界环境的急剧影响时,会改变所表达的基因,产生因子更替的现象。 • 如环境温度升高时,大肠杆菌会开启rpoH基因的表达, 其产物32能识别热激基因的启动子,而正常状态下表达的基因会关闭或表达水平下降。 • 芽孢菌生活周期过程中生活方式的改变也是通过因子的更替完成的。
第一节 转录酶1 原核生物的RNA聚合酶2 真核生物的RNA聚合酶
2.1 真核RNA聚合酶的结构特点: 1)大分子蛋白质,500KDa或更多; 2) 分子量500KDa, 含两个大亚基和 7~12 个小亚基。 RNApolⅡ: 3) 大亚基中有 羧端功能区(carboxy terminal domain CTD) CTD中含一保守氨基酸序列的多个重复 Tyr-Ser-Pro-Thr-Ser-Pro-Ser C 端重复七肽 不同生物中重复数目不一样(酶活性)
CTD: • 参与转录起始 • 真核RNA pol独有。 • CTD参与转录 → ⅡB → ⅡA → 使 RNApol易于离开启动子进入延伸过程(10倍)
2.2 线粒体和叶绿体的RNA pol • RNA pol较小,更类似于细菌的RNA pol。 • RNA pol转录控制也很简单,只转录少数几个基因 。
第一节 转录酶 • 第二节 启动子 • 第三节 终止子 • 第四节 转录的机制 • 第五节 转录产物的后加工 • 第六节 RNA的剪接
第二节 启动子 1 原核生物的启动子 2 真核生物的启动子
1 原核生物的启动子 启动子(promoter):DNA分子与RNA聚合酶特异结合的部位,也就是转录开始的部位。 • 不同启动子对RNA聚合酶的亲和力各不同, • 根据其合成蛋白质的多少区别为强弱启动子。 • 强启动子与RNA聚合酶亲和性高,合成的mRNA多,翻译的蛋白质也多。
-10 +1 +10 upstream start point downstream • DNA上开始转录的第一个碱基定为+1, • 沿转录方向顺流而下的核苷酸序列均用正值表示; • 逆流而上的核苷酸序列均用负值表示。
典型原核生物启动子的结构 1.1 原核生物启动子的结构特点 -35 -10 转录起点 TTGACA 16-19bp TATAAT 5-9bp
1) 转录起始点 1.1 原核生物启动子的结构特点 • 多数情况下(>90%)为嘌呤, • 常见序列为CAT,A为起始点
2) -10区 又称为Pribnow盒 • 结构特点: • 保守序列:TATAAT(T80A95T45A60A50T96) • A.T较丰富,易于解链。 • 功能: (1) RNA pol结合位点 ; (2) 形成开放启动复合体; (3) 使RNA pol定向转录。
3) -35区 又称为Sextama box 结构特点: • 其保守序列 TTGACA(T82T84G78A65C54A45) • 与-10序列,相隔16-19bp。 功能: • (1) 为RNA pol的识别位点。 • σ亚基识别-35序列,为转录选择模板链。 • RNA Pol 核心酶和模板结合,进行聚合; (2) 很大程度上决定了启动子的强度。
-35 序列与-10 序列与转录效率的关系 标准启动子 -35 TTGACA -10 TATAAT 不同的启动子 a. 与标准启动子序列同源性越高 →启动强度越大 b. 与标准启动子同源性越低 →启动强度越小 c. 与标准启动子差异很大时 →由另一种σ因子启动
4) -10和-35之间距离: • 间距非常重要,16-19bp的间距转录效率最高,碱基序列并不重要 • 间距上的突变种类: 间距趋向于17bp → 上升突变 间距远离17bp → 下降突变 原因: • 该距离大致是双螺旋绕两圈的长度。这两个结合区是在DNA分子的同一侧面,此酶结合在双螺旋的一面。 • 原核生物亦有少数启动子缺乏这两个序列之一。RNA聚合酶往往不能单独识别这种启动子,需要有辅助蛋白质的帮助。
5)启动子附近其他DNA序列: • 起始位点上游50到150bp之间的序列是启动子的完全活性所必需的。 • 上游序列可以吸引拓扑异构酶,后者可导致结合的局部产生有利于转录起始的超螺旋状态。 • 远离部位序列富有AT,能增进转录起始的频率。
保守序列 大肠杆菌RNA聚合酶全酶所识别的启动子区 T80A95T45A60T96 T82T84G78A65C54A45
第二节 启动子 1 原核生物的启动子 2 真核生物的启动子
2 真核生物的启动子 2.1 RNApolⅡ的启动子 2.2 RNApol I的启动子 2.3 RNApol III的启动子 三种 RNApol → 三种转录方式 三种启动子 → 三类基因,Ⅰ类 Ⅱ类 Ⅲ类
2.1 RNApolⅡ的启动子 • 通用型启动子(无组织特异性) • 结构最复杂 • 位于转录起始点的上游 • 有多个短序列元件组成
(1) 帽子位点(cap site):转录起始位点 与Prok.的相似 ,多为A (2) TATA框(Hogness 框或 Goldberg-Hogness 框) 结构特点: • 位于-30处 • 一致序列为T82A97T93A85A63(T37)A83A50(T37) 功能: • 定位转录起始点 (类似原核的Pribnow框), 也称为选择子(selector) • TATA是绝大多数真核基因的正确表达所必需的
(3) CAAT框(CAAT box) 结构特点: • 位于-75bp处 • 一致序列为GGC/TCAATCT 功能: • 前两个 G 的作用十分重要(转录效率) • 增强启动子的效率、频率,不影响启动子的特异性 (距转录起始点的距离,正反方向) ☻ 以上三个保守序列在绝大多数启动子中都存在
(4) GC框 (GC box) • 位于-90附近,较常见的成分 • 核心序列为GGGCGG • 可有多个拷贝,也可以正反两方向排列 (5)其他元件 • 八聚核苷酸元件(octamer element OCT元件) 一致序列为 ATTTGCAT • KB元件 一致序列为 GGGACTTTCC • ATF元件 一致序列为 GTGACGT • 以及还有一些位于起始点下游的相关元件
(6) 起始子(initiator,Inr): • 位于起始点 -3~+5 • Py2CAPy5构成, • 可能提供RNA pol Ⅱ识别。 • 当有的基因缺少TATA框时,可能由Inr来替代它的作用, • 无论TATA是否存在,Inr对于启动子的强度和起始位点的选择都是十分重要的 。
小结--- 与原核生物启动子的比较: (1)多种元件:TATA框,GC框,CAAT框,OCT等; (2)不同元件的组合情况:位置、序列、距离和方向都不完全相同; (3)需转录因子参与转录的全过程:转录因子先和启动子结合,再与RNA聚合酶一起形成转录起始复合物,开始转录的过程。
真核启动子含有不同的组件 SV40 早期启动子 胸苷激酶 组蛋白H2B -140 -120 -100 -80 -60 -40 -20 +1 Oct CAAT GC TATA
