Download
1 / 1
10 likes | 149 Views
Epidemiology of non-B clade forms of HIV-1 in MSM in the UK. Fox J 1 , Duda A 2 ,Green H 3 , Dunn D 3 , Kaye S 1 , McClure M 1 , Fidler S 1

E N D
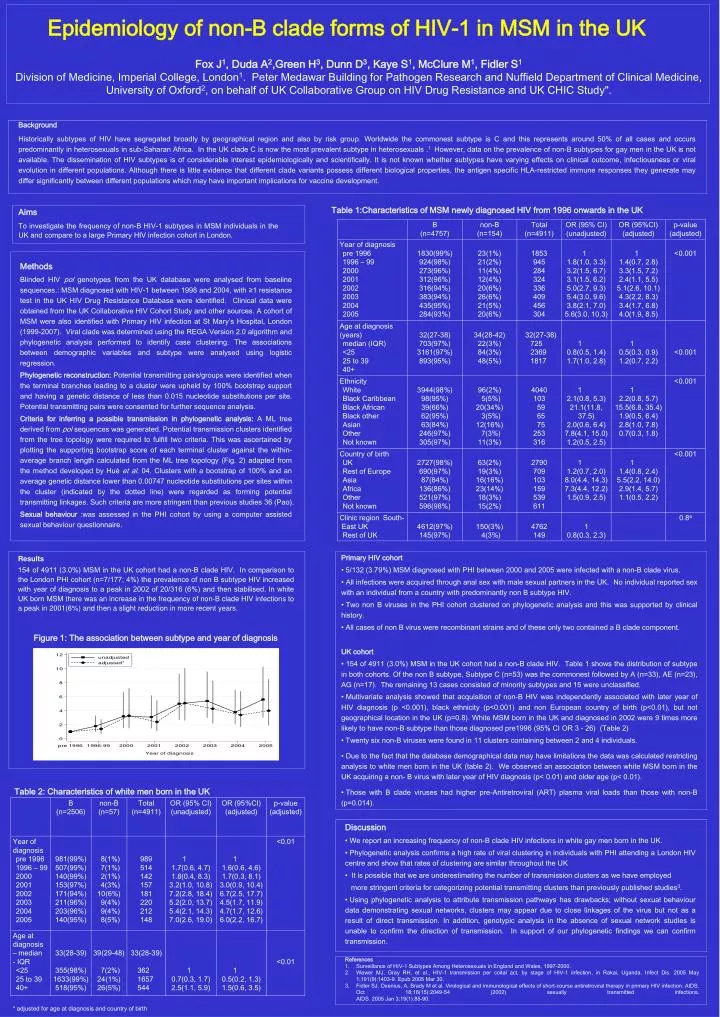
Epidemiology of non-B clade forms of HIV-1 in MSM in the UK Fox J1, Duda A2,Green H3, Dunn D3, Kaye S1, McClure M1, Fidler S1 Division of Medicine, Imperial College, London1. Peter Medawar Building for Pathogen Research and Nuffield Department of Clinical Medicine, University of Oxford2, on behalf of UK Collaborative Group on HIV Drug Resistance and UK CHIC Study". Background Historically subtypes of HIV have segregated broadly by geographical region and also by risk group. Worldwide the commonest subtype is C and this represents around 50% of all cases and occurs predominantly in heterosexuals in sub-Saharan Africa. In the UK clade C is now the most prevalent subtype in heterosexuals .1 However, data on the prevalence of non-B subtypes for gay men in the UK is not available. The dissemination of HIV subtypes is of considerable interest epidemiologically and scientifically. It is not known whether subtypes have varying effects on clinical outcome, infectiousness or viral evolution in different populations. Although there is little evidence that different clade variants possess different biological properties, the antigen specific HLA-restricted immune responses they generate may differ significantly between different populations which may have important implications for vaccine development. Table 1:Characteristics of MSM newly diagnosed HIV from 1996 onwards in the UK Aims To investigate the frequency of non-B HIV-1 subtypes in MSM individuals in the UK and compare to a large Primary HIV infection cohort in London. Methods Blinded HIV pol genotypes from the UK database were analysed from baseline sequences.: MSM diagnosed with HIV-1 between 1998 and 2004, with ≥1 resistance test in the UK HIV Drug Resistance Database were identified. Clinical data were obtained from the UK Collaborative HIV Cohort Study and other sources. A cohort of MSM were also identified with Primary HIV infection at St Mary’s Hospital, London (1999-2007). Viral clade was determined using the REGA Version 2.0 algorithm and phylogenetic analysis performed to identify case clustering. The associations between demographic variables and subtype were analysed using logistic regression. Phylogenetic reconstruction:Potential transmitting pairs/groups were identified when the terminal branches leading to a cluster were upheld by 100% bootstrap support and having a genetic distance of less than 0.015 nucleotide substitutions per site. Potential transmitting pairs were consented for further sequence analysis. Criteria for inferring a possible transmission in phylogenetic analysis: A ML tree derived from pol sequences was generated. Potential transmission clusters identified from the tree topology were required to fulfill two criteria. This was ascertained by plotting the supporting bootstrap score of each terminal cluster against the within-average branch length calculated from the ML tree topology (Fig. 2) adapted from the method developed by Huéet al. 04. Clusters with a bootstrap of 100% and an average genetic distance lower than 0.00747 nucleotide substitutions per sites within the cluster (indicated by the dotted line) were regarded as forming potential transmitting linkages. Such criteria are more stringent than previous studies 36 (Pao). Sexual behaviour :was assessed in the PHI cohort by using a computer assisted sexual behaviour questionnaire. • Primary HIV cohort • 5/132 (3.79%) MSM diagnosed with PHI between 2000 and 2005 were infected with a non-B clade virus. • All infections were acquired through anal sex with male sexual partners in the UK. No individual reported sex with an individual from a country with predominantly non B subtype HIV. • Two non B viruses in the PHI cohort clustered on phylogenetic analysis and this was supported by clinical history. • All cases of non B virus were recombinant strains and of these only two contained a B clade component. • UK cohort • 154 of 4911 (3.0%) MSM in the UK cohort had a non-B clade HIV. Table 1 shows the distribution of subtype in both cohorts. Of the non B subtype, Subtype C (n=53) was the commonest followed by A (n=33), AE (n=23), AG (n=17). The remaining 13 cases consisted of minority subtypes and 15 were unclassified. • Multivariate analysis showed that acquisition of non-B HIV was independently associated with later year of HIV diagnosis (p <0.001), black ethnicity (p<0.001) and non European country of birth (p<0.01), but not geographical location in the UK (p=0.8). White MSM born in the UK and diagnosed in 2002 were 9 times more likely to have non-B subtype than those diagnosed pre1996 (95% CI OR 3 - 26) (Table 2) • Twenty six non-B viruses were found in 11 clusters containing between 2 and 4 individuals. • Due to the fact that the database demographical data may have limitations the data was calculated restricting analysis to white men born in the UK (table 2). We observed an association between white MSM born in the UK acquiring a non- B virus with later year of HIV diagnosis (p< 0.01) and older age (p< 0.01). • Those with B clade viruses had higher pre-Antiretroviral (ART) plasma viral loads than those with non-B (p=0.014). Results 154 of 4911 (3.0%)MSM in the UK cohort had a non-B clade HIV.In comparison to the London PHI cohort (n=7/177; 4%) the prevalence of non B subtype HIV increased with year of diagnosis to a peak in 2002 of 20/316 (6%) and then stabilised. In white UK born MSM there was an increase in the frequency of non-B clade HIV infections to a peak in 2001(6%) and then a slight reduction in more recent years. Figure 1: The association between subtype and year of diagnosis Table 2: Characteristics of white men born in the UK • Discussion • We report an increasing frequency of non-B clade HIV infections in white gay men born in the UK. • Phylogenetic analysis confirms a high rate of viral clustering in individuals with PHI attending a London HIV centre and show that rates of clustering are similar throughout the UK • It is possible that we are underestimating the number of transmission clusters as we have employed • more stringent criteria for categorizing potential transmitting clusters than previously published studies3. • Using phylogenetic analysis to attribute transmission pathways has drawbacks; without sexual behaviour data demonstrating sexual networks, clusters may appear due to close linkages of the virus but not as a result of direct transmission. In addition, genotypic analysis in the absence of sexual network studies is unable to confirm the direction of transmission. In support of our phylogenetic findings we can confirm transmission. • References • Surveillance of HIV-1 Subtypes Among Heterosexuals in England and Wales, 1997-2000. • Wawer MJ, Gray RH, et al., HIV-1 transmission per coital act, by stage of HIV-1 infection, in Rakai, Uganda. Infect Dis. 2005 May 1;191(9):1403-9. Epub 2005 Mar 30. • Fidler SJ, Oxenius, A, Brady M et al. Virological and immunological effects of short-course antiretroviral therapy in primary HIV infection. AIDS. Oct 18;16(15):2049-54 (2002) sexually transmitted infections.AIDS. 2005 Jan 3;19(1):85-90. * adjusted for age at diagnosis and country of birth






















