Download
1 / 1
10 likes | 118 Views
High-Resolution, Accurate-Mass (HR/AM) a nd Intelligent Acquisition-Enabled Global Discovery and Quantification of Histones, Histone PTMS, and Histone Modification Enzymes in Mesenchymal Stem Cells

E N D
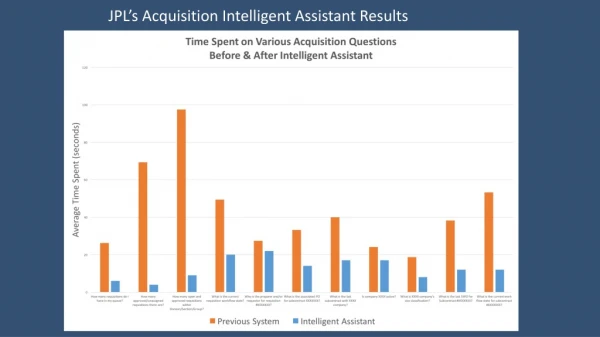
High-Resolution, Accurate-Mass (HR/AM) and Intelligent Acquisition-Enabled Global Discovery and Quantification of Histones, Histone PTMS, and Histone Modification Enzymes in Mesenchymal Stem Cells Amol Prakash1, Maryann Vogelsang1, David Sarracino1, Scott Peterman1, Victoria Lunyak2, Benny Blackwell2, James Tollervey2, Shadab Ahmad1, Gregory Byram1, Bryan Krastins1, Mary Lopez11BRIMS, Thermo Fisher Scientific, Cambridge, MA;2Buck Institute for Age Research, Novato, CA FIGURE 5 Differential abundance of cytoplasmic fraction histones and histone modification proteins in mesenchymal stem cells with acute DNA damage (2 hr treatment with bleomycin) and DNA damage-induced cellular senescence (cellular recovery after 5 days of post-bleomycin treatment). FIGURE 3. Differential abundance of nuclear fraction histones and histone modification proteins in mesenchymal stem cells with acute DNA damage (2 hr treatment with bleomycin) and DNA damage-induced cellular senescence (cellular recovery after 5 days of post-bleomycin treatment). Results Intelligent Data Acquisition Initial discovery experiments were performed to help drive targeted quantitative experiments (Figures 1, 2). The discovery experiments were performed in an unbiased data-dependent acquisition across different biological samples to determine proteins and peptides as well as the initial set of PTMs. From the initial discovery results, histones and histone modification proteins were chosen and further targeted discovery experiments were performed to increase the probability of modified peptide identification based on theoretical m/z inclusion lists. The combined results from the discovery experiments were used to build a local spectral library consisting of precursor and product ion m/z values and relative abundance distribution as well as relative retention time values. A set of peptides from the discovery data was selected based on known and novel PTMs. The spectral library information for the targeted peptides was used to create a targeted inclusion list and reference information to perform qual/quan determination in real time. The real-time feedback facilitated optimization of instrument parameters to maximize instrument duty cycle and detection capabilities resulting in significantly increasing the number of peptides quantified per experiment. The final assay performed qual/quan studies on 36 proteins and 154 histone and histone-related peptides and modified analogs (Table 1). The combined approach enabled quantification of previously identified modified peptides as well as novel targets across different samples and were correlated to somatic or stem cell aging (replicative or genotoxic stress-induced senescence) (Figs 3-5). TABLE 1. Targeted proteins and peptides Overview Purpose: Development of a real-time, intelligent acquisition strategy for HR/AM global targeted quantification of histones, histone PTMs, and histone modification enzymes Methods: Mesenchymal stem cells were derived from human adipose tissue and treated with bleomycin.Samples were analyzed on a Thermo Scientific™ Q Exactive™ mass spectrometer using an intelligent acquisition method. Results: The final assay performed qual/quan studies on 36 proteins and 154 histone and histone-related peptides and modified analogs. The combined approach enabled quantification of previously identified modified peptides as well as novel targets across different samples and were correlated to somatic or stem cell aging (replicative or genotoxic stress-induced senescence). Introduction Chromatin is viewed as an operational interface for almost all known nuclear processes. Nucleosomal packaging and histone modifications dictate the different degrees of primary chromatin compaction achieved by additional chromatin structural proteins. For example, euchromatic chromatin fibers contain six nucleosomes per 11 nm; however, heterochromatin consists of 12–15 nucleosomes per 11 nm. A dynamic balance between these two radically different chromatin compaction states is at the very core of the high-level nuclear chromatin organization (nuclear architecture), and is vital for maintaining cell-type identity over time. Functional differences between the cells in an organism are defined by epigenetic factors and epigenetic programs, which are critical for the preservation of functional integrity of the cellular phenotypes (1). However, as a mediator of the external signals, chromatin is anything but static. Nucleosome unwrapping and disassembly events, which must occur during DNA replication, transcription, and DNA repair, can directly influence the state of chromatin compaction. Several lines of evidence obtained in Drosophila, yeast, and plants indicate that chromatin undergoes disassembly during the onset of DNA double-strand breaks (DSB) and repair at the DSB sites. It is still an open question how to quantify the chromatin changes. One of the accepted ways to do such is through cellular fractionation. Based on the salt and detergent concentrations in buffer solutions, three cellular fractions can be obtained: cytoplasmic, nuclear soluble (proteins that are loosely bound to chromatin), and a chromatin fraction (containing tightly bound proteins, or enzymes engaged in chromatin modifications). Tracing histones or histone-modifying activities (HMT, HATs, and HDACs) dynamics in these fractions during biological events permits the assessment of overall chromatin dynamics and full characterization of the cellular stage (2). In this study, we set out to quantify the changes in the chromatin composition in primary human stem cells upon acute DNA damage and during drug-evoked senescence. This assessment allowed insights into the “access, repair, restore” model of chromatin dynamics upon DNA damage repair and a better understanding of whether or not misregulation of this axis is one of the critical factors of cellular senescence. In our model system, we compared two conditions: 1) Acute DNA damage (2 hrs treatment with bleomycin) and 2) DNA damage-induced cellular senescence (cellular recovery after 5 days of post-bleomycin treatment). Both conditions were compared to the normally proliferating (self-renewing) cells. Using a novel application of intelligent acquisition and HR/AM MS, we have developed workflows for quantitative global profiling and targeted analysis of histones, histone post-translational modifications (PTMs), and histone modification enzymes. Methods Samples Mesenchymal stem cells were derived from human adipose tissue and treated with bleomycin as follows: nuclear, cytoplasmic, and chromatin fraction samples were isolated from proliferating (bleomycin –) cells as well as cells under conditions of acute DNA damage (bleomycin 2 hrs) and drug-induced senescence (bleomycin 2 hrs followed by a 5-day recovery). MS Data Acquisition and Analysis Samples were analyzed on a Q Exactive mass spectrometer. Initial discovery experiments were performed to generate a list of peptides/protein IDs and corresponding spectral libraries. A subset of peptides was selected for targeted quantification across the different samples. Heavy labeled analogs were used for qual/quan data processing. Qualitative and quantitative data analysis was performed using Thermo Scientific™ Pinpoint™ software. A B C D FIGURE 1. Intelligent data acquisition strategy. Pictorial representation of intelligent data acquisition schemes for targeted peptide quantification using a targeted scanning window, target elution identification, and real-time product ion spectral acquisition. Both precursor and product ion spectral matching are performed to increase the selectivity of data acquisition. • Conclusion • We have developed a real-time, intelligent acquisition strategy for HR/AM global targeted quantification of histones, histone PTMs, and histone modification enzymes in primary human stem cells upon acute DNA damage and during drug-evoked senescence. • Assessment of the histone H1, H2a, H2B, H4, and H3 family member abundance in the soluble nuclear fraction of the cells subjected to genotoxic drug-induced senescence (5 days after exposure to bleomycin), demonstrated that the dynamics of histone binding in the senescent cells changes significantly (Figure 3B). Our data also suggest that the composition of the nucleosomal particles undergoes dramatic changes upon senescence. We have observed significant reduction in chromatin-bound histone H4 and all of the members of the H1 family (Figure 3B). These data suggest that with reduction of H1 histone family members in the chromatin of senescent cells, the overall compaction of chromatin fiber decreases dramatically. • The representation of macro H2A histone does not change with senescent-specific transformation. Levels of H2B type1-A and type1-B histones also do not change significantly. These data suggest histone type specificity in chromatin dynamics upon reaching a senescent state. • A dramatic increase in chromatin-bound HAT p300 and HMT MLL3 correlates well with the loss of histone 1 family members from the chromatin. These data support the hypothesis that relaxation of chromatin compaction in senescent cells might be a leading cause of increased transcriptional noise (transcriptional leakage) upon senescence. • References • Lunyak, V.V.; Tollervey, J.R. Epigenetics: judge, jury and executioner of stem cell fate. Epigenetics. 2012 Aug;7(8):823–40. doi: 10.4161/epi.21141. Epub 2012 Jul 18. • Lopez, M.F.; Tollervey, J.; Krastins, B.; Garces, A.; Sarracino, D.; Prakash, A.; Vogelsang, M.; Geesman, G.; Valderrama, A.; Jordan, I.K.; Lunyak, V.V. Depletion of nuclear histone H2A variants is associated with chronic DNA damage signaling upon drug-evoked senescence of human somatic cells. Aging (Albany NY). 2012 Nov;4(11):823–42. Most intense isotope 2nd most intense isotope FIGURE 4. Differential abundance of chromatin associated histones and histone modification proteins in mesenchymal stem cells with acute DNA damage (2 hr treatment with bleomycin) and DNA damage-induced cellular senescence (cellular recovery after 5 days of post-bleomycin treatment). A B FIGURE 2. Extracted ion chromatograms of fragment ions from KESYSIYVKYK B. Precursor ion charge state 2 A. Precursor ion charge state 3 C D All trademarks are the property of Thermo Fisher Scientific and its subsidiaries. This information is not intended to encourage use of these products in any manners that might infringe the intellectual property rights of others.