Download
1 / 28
280 likes | 297 Views
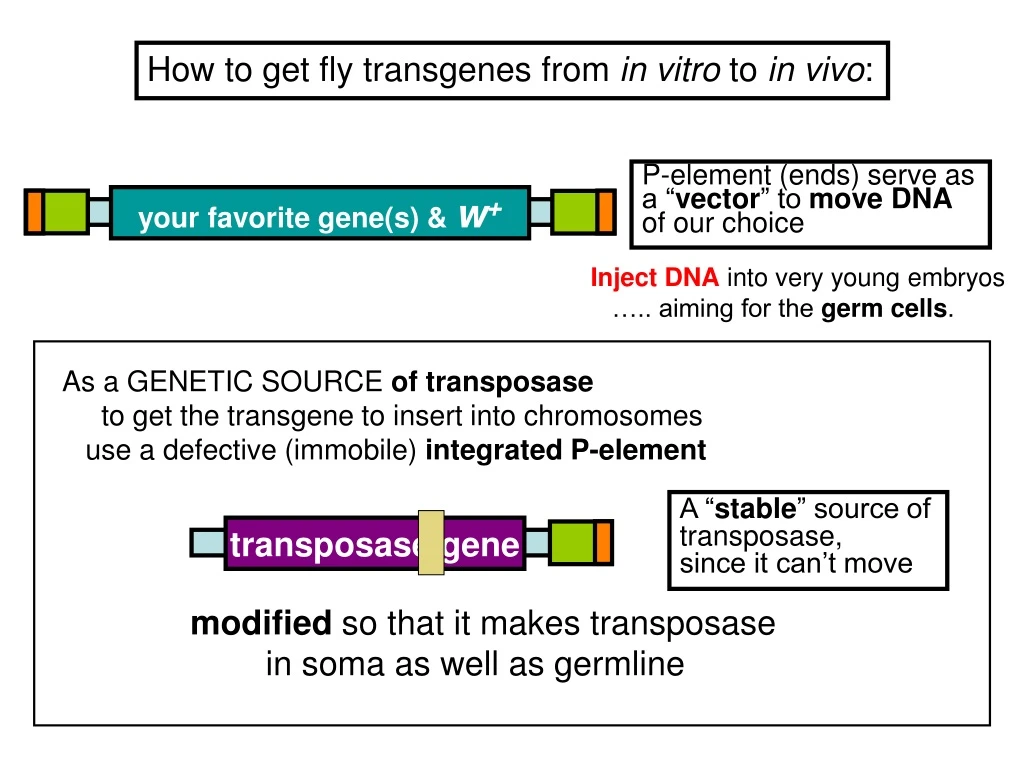
P-element (ends) serve as a “ vector ” to move DNA of our choice. Inject DNA into very young embryos ….. aiming for the germ cells. As a GENETIC SOURCE of transposase to get the transgene to insert into chromosomes use a defective (immobile) integrated P-element.

E N D
P-element (ends) serve as a “vector” to moveDNA of our choice Inject DNA into very young embryos ….. aiming for the germ cells. As a GENETIC SOURCE of transposase to get the transgene to insert into chromosomes use a defective (immobile) integrated P-element A “stable” source of transposase, since it can’t move transposase gene your favorite gene(s) & w+ modified so that it makes transposase in soma as well as germline How to get fly transgenes from in vitro to in vivo:
w+ 2 whatever is useful (eg. w+) Use the engineered mobile genetic element as a mutagen (make it hop randomly into genes): (a nonautonomous element) starting element: It is a "genetically tagged" mutagen (we can follow it)
transposase to mobilize (pseudo-replicative transposition) Tp X w+ new (random) site of insertion 2 w+ w+ (not carrying the transposase source) X collect progeny from the appropriate genetic cross of the dysgenic parent 3 (not carrying the original P) 2 3 gene starting element: happening In the dysgenic parent's germline If so, already well marked and easy to clone! Is this a mutant allele of interest?
your favorite gene(s) & w+ P P (a nonautonomous element) If we are going to want to use as a mutagen (hop into genes)… Lucky thing that M strains exist (strains with no pre-existing source of P transposase or antitransposase to interfere with our controlling non-autonomous element [transgene] mobility)
Why do M strains exist? Lab strains taken from the wild before 1950’s don’t have P elements. This DNA parasite invaded D. melanogaster sometime in the 1950s, then rapidly spread nearest P-element relative: in a fly species 50 Myr diverged -- an example of horizontal genetic transmission How? (xfrd in same generation, i.e. gene not introduced from parents (vs. vertical) Are fly-workers to blame for contaminating D. melanogaster?
tagged transposon hopped in the dysgenic parent's germline collect progeny from the dysgenic parent w+ 3 gene new (random) site of insertion How do we know that we have even mutated a gene, much less generated a mutant allele of use/interest ? recover the chromosome in a MUTANT SCREEN or SELECTION
Steps in forward genetics: decide what to study generate informative mutant alleles (mutagenesis) recover informative mutant alleles study informative mutant allele (do molecular biology) write paper reap rewards
Nobel Prize 1995 NATURE V287:p795 (1980) (products of a mutant screen)
wildtype gooseberry patch wildtype gooseberry “skins” of dead larvae patch mutant phenotypes informative for understanding pattern formation in metazoans
text: 20.3 “The genetic analysis of body-plan development in Drosophila: a comprehensive example. (pp732-745)
They regulate… Drosophila homeotic gene clusters the segmentation regulatory gene hierarchy Ant. Pst. MammalianHox gene clusters maternal effect gap Mouse embryo pair- rule segment polarity Fig. 20.22 p741 Fig. 20.26 p744
Two general categories of mutant allele recovery strategies: (“brute force” screens) (1) geneticscreens make mutations randomly, then you sift through chromosomes (often one at a time) looking for mutant alleles of interest/use (2) genetic selections make mutations randomly, then let nature eliminate all undesired mutant alleles so you are only left with the good stuff 2 easier to execute than 1, butoften not possible to design & potentially more biased (may get only what you think to look for)
generate mutant allele generate mutant allele recover mutant allele recover mutant allele study mutant allele study mutant allele maintain mutant allele write paper write paper reap rewards reap rewards By the way: an important but under-appreciated step in genetic analysis: for each fly line: transfer to 15ml new food every 3 weeks Homework problem: How much food (corn meal, molasses, yeast) has T.H.Morgan’s original white1 mutant line consumed since 1910? Strategies & tools that help us recover mutant alleles can also help us maintain them.
To freeze or not to freeze seeds Maintaining mutant stocks (lines) in model genetic systems: mouse most microbes (embryos) (spores are nice) fish arabidopsis (“the weed”) & corn fly worm
Hence: screen/selection schemes must provide for the recovery of recessive lethals and steriles Basic facts to consider in designing screens and selections: (1) Most LOF mutant alleles are recessive (all else being equal) (LOF mutations are the most frequent class) (2) Most null alleles of genes with an obvious LOF phenotype are lethal, or at least sterile. (3) Most “developmentally interesting” genes are essential for viability or fertility
Diploid: lethal/ + alive (fertile) Haploid: lethal dead (sterile) often for microbes rely on conditional lethals in generating mutations: The “diploid advantage” for recessive lethal studies: (+ holds the fort) (let naked and exposed)
condition A condition B growth vs. no growth augmented by: p547: Fig. 15.5 (Penicillin) enrichment Haploid: lethal dead (sterile) rely on conditional lethals in generating mutations: genetic screen mutant screen or selection? all grow (mutagenize wildtype) onlymutants of interest don’t grow Two key tricks for microbes: p212: Fig. 7.5 Replica Plating & p558: Fig. 15.15 genetic selection
condition A condition B growth vs. no growth all grow (mutagenized) only mutants of interest don’t grow Two key tricks for microbes: p212: Fig. 7.5 & its use: Fig. 15.15 (p558) Replica Plating genetic screen
genetic selection (diluting out the penicillin) Replica plate on condition A condition B growth vs. no growth augmented by: p547: Fig. 15.5 (Penicillin) enrichment all grow (mutagenized) only mutants of interest don’t grow
The “diploid advantage” for recessive lethal studies: The “diploid handicap” for recessive lethal studies: Diploid: + masks lethal lethal/ + alive (fertile) (+ holds the fort) Haploid: effects of lethal immediately obvious lethal dead (sterile) (let naked and exposed)
mutagenize Male Female :PARENTS form zygotes X +/+ +/+ + + + + + + + + + + + + + + + + + + + + + d a a +/+ +/ b b a c +/+ eggs +/ b sperm PROGENY The problem with diploids in hunting for new recessive mutations: F1
mutagenize Male Female X +/+ +/+ +/ a +/ b The problem with diploids in hunting for new recessive mutations: +/+ +/+ F1 PROGENY given: we are interested in (finding) the a-/a- phenotype How do we know who (if anyone) is carrying a-? …the individual who can produce a-/a- offspring.
mutagenize Male Female X +/+ +/+ If we can “self” this individual, we are effectively mating to +/a- for sure +/ a +/ b YES! & we know in the F2 of course, we had to self everyone: Noa-/a- a-/a- The problem with diploids in hunting for new recessive mutations: Which is the individual who can produce a-/a- offspring? +/+ +/+ F1 PROGENY To whom do we mate to find out?
Male Female X +/+ +/+ +/+ +/+ +/ +/ +/ a a a +/+ +/ b Meanwhile +/+ +/+ +/+ +/+ +/+ +/+ +/+ The problem with diploids in hunting for new recessive mutations: mutagenize F1 +/+ +/+ To whom do we mate to find out -- if we can’t self? …. we still don’t know in the F2!
mutagenize Male Female X +/+ +/+ ..but at least now we have potential mates with a- +/ +/ +/ male? a a a +/+ +/ b Mate inter se +/+ +/+ +/+ +/+ +/+ +/+ female? +/+ Mate inter se a /a at least some chance: +/+ at best The problem with diploids in hunting for new recessive mutations: -- if we can’t self. …. we still don’t know in the F2! F1 +/+ +/+ X+/+ X+/+ (must keep populations separate!) F2
Male Female X +/+ +/+ +/+ +/ +/ +/ a a a +/ b +/+ +/+ +/+ +/+ +/+ if we cross them, a-/a- will come: a /a +/+ only The problem with diploids in hunting for new recessive mutations: -- if we can’t self. mutagenize F1 +/+ +/+ X F2 +/+ can we do better than mating inter se? +/+ +/+ Mate inter se
mutagenize Male Female X += non-mutagenized from original Mom +/+ +/+ F1 +/+ +/+ +/+ X F2 +/+ +/ +/ +/ a a a +/+ +/ +/+ b if we cross them, a-/a- will come: a /a It would help if we could keep track of chromosomes: a-= mutagenized chromosome with new mut. += mutagenized but not desired mutant += non-mutagenized from F1 mate Even nicer if we could eliminate extraneous animals we can do better than mating inter se
our friend, Herman Muller had the answer (early ‘30s): (1)used them to determine mutation frequency: …how often a new recessive lethal arose on a given fly chromosome (2) used them to “maintain” deleterious recessive alleles of interest Balancer chromosomes: (a) a chromosome you can distinguish from the others. dominant marker mutant alleles (Bar, Curly, Stubble) (b) a chromosome that will notrecombine with others crossover suppressors (multiple inversions) (c) a chromosome that will not “become” homozygous (i.e. that would either be lethal or sterile if homozygous) recessive lethal or sterile alleles