Download
1 / 1
10 likes | 98 Views
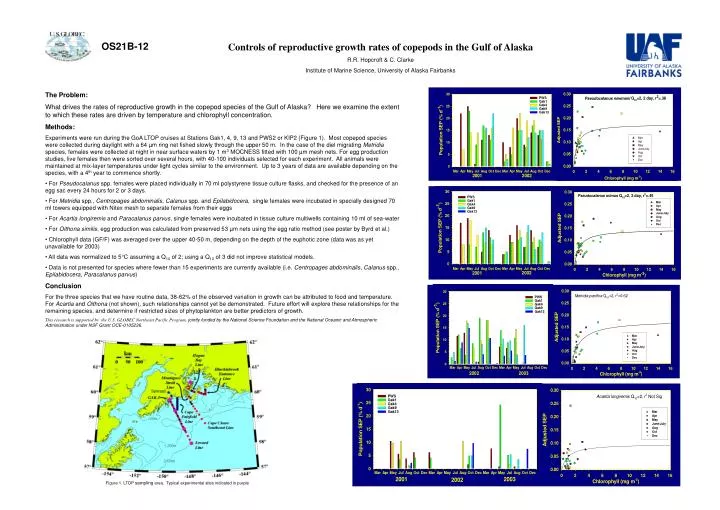
OS21B-12. Controls of reproductive growth rates of copepods in the Gulf of Alaska R.R. Hopcroft & C. Clarke Institute of Marine Science, University of Alaska Fairbanks. The Problem:

E N D
OS21B-12 Controls of reproductive growth rates of copepods in the Gulf of Alaska R.R. Hopcroft & C. Clarke Institute of Marine Science, University of Alaska Fairbanks • The Problem: • What drives the rates of reproductive growth in the copepod species of the Gulf of Alaska? Here we examine the extent to which these rates are driven by temperature and chlorophyll concentration. • Methods: • Experiments were run during the GoA LTOP cruises at Stations Gak1, 4, 9, 13 and PWS2 or KIP2 (Figure 1). Most copepod species were collected during daylight with a 64 µm ring net fished slowly through the upper 50 m. In the case of the diel migrating Metridia species, females were collected at night in near surface waters by 1 m3 MOCNESS fitted with 100 µm mesh nets. For egg production studies, live females then were sorted over several hours, with 40-100 individuals selected for each experiment. All animals were maintained at mix-layer temperatures under light cycles similar to the environment. Up to 3 years of data are available depending on the species, with a 4th year to commence shortly. • For Pseudocalanus spp. females were placed individually in 70 ml polystyrene tissue culture flasks, and checked for the presence of an egg sac every 24 hours for 2 or 3 days. • For Metridia spp., Centropages abdominalis, Calanus spp. and Epilabidocera, single females were incubated in specially designed 70 ml towers equipped with Nitex mesh to separate females from their eggs • For Acartia longiremis and Paracalanus parvus, single females were incubated in tissue culture multiwells containing 10 ml of sea-water • For Oithona similis, egg production was calculated from preserved 53 µm nets using the egg ratio method (see poster by Byrd et al.) • Chlorophyll data (GF/F) was averaged over the upper 40-50 m, depending on the depth of the euphotic zone (data was as yet unavailable for 2003) • All data was normalized to 5°C assuming a Q10 of 2; using a Q10 of 3 did not improve statistical models. • Data is not presented for species where fewer than 15 experiments are currently available (i.e. Centropages abdominalis, Calanus spp., Epilabidocera, Paracalanus parvus) • Conclusion • For the three species that we have routine data, 38-62% of the observed variation in growth can be attributed to food and temperature. For Acartia and Oithona (not shown), such relationships cannot yet be demonstrated. Future effort will explore these relationships for the remaining species, and determine if restricted sizes of phytoplankton are better predictors of growth. • This research is supported by the U.S. GLOBEC Northeast Pacific Program, jointly funded by the National Science Foundation and the National Oceanic and Atmospheric Administration under NSF Grant OCE-0105236. Figure 1. LTOP sampling area. Typical experimental sites indicated in purple






![Chapter 7: Computer-Assisted Audit Techniques [CAATs]](https://cdn0.slideserve.com/534186/chapter-7-computer-assisted-audit-techniques-caats-dt.jpg)