Download
1 / 1
10 likes | 190 Views
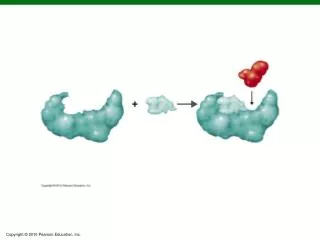
3D structure of β -lactoglobulin A. The hydrophobic cavity region is defined by five β -strands on the front (strands A, H, G, F, E) and three β -strands on the back (strands B, C and D). In the closed conformation (pH=2), E-F loop covers the hydrophobic cavity opening.

E N D
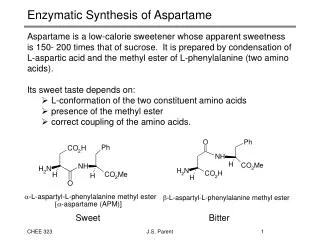
3D structure of β-lactoglobulin A • The hydrophobic cavity region is defined by five β-strands on the front (strands A, H, G, F, E) and three β-strands on the back (strands B, C and D). • In the closed conformation (pH=2), E-F loop covers the hydrophobic cavity opening. • The “α-helix” between strands H and I is also thought to be associated with the hydrophobic cavity region. • In the open conformation (pH>7), the E-F loop moves, exposing the residues within the hydrophobic cavity and also changing the conformation of the α-helix. (Uhrinova et.al., Biochemistry (2000) 39,3565) Hydrophobic cavity region http://www.rcsb.org/pdb/cgi/resultBrowser.cgi RCSB PDB entry 1CJ5 Enzymatic Cleavage of B-lactoglobulin A LC/MS spectrum of the peptides at tr = 38.94 LC-MS-MS spectrum of the doubly charged parent ion at m/z 855.1 Protein Surface Mapping by Mass Spectrometry to Monitor pH-Dependent Structural Transitions in Beta-lactoglobulin A* Robert L. Hettich 1; Demet Ataman1,2; Joshua Sharp 3 1Oak Ridge National Laboratory, Oak Ridge ,TN; 2 ORNL-UTK Genome Science and Technology Graduate School, Oak Ridge, TN, 3 NIEHS, Research Triangle Park, NC OVERVIEW • Information about protein conformation is important for determining biological function • One method of probing higher order protein structure is chemical tagging of the solvent accessible residues using oxidation as a labeling technique (Maleknia and Chance, Anal. Chem. (1999) 71, 3965) • We have demonstrated a protein surface mapping technique based on hydroxyl radical attachment with high-resolution mass spectrometry identification to probe the solvent accessibility of the protein’s native conformation (Sharp, et.al. Anal. Biochem. (2003) 313, 216) • This surface mapping technique was used to investigate the pH-dependent conformational changes of β-lactoglobulin A, providing specific details of the opening of the hydrophobic cavity RESULTS FOR β-LACTOGLOBULIN A OXIDATION AFTER TRYPTIC DIGESTION RESULTS FOR β-LACTOGLOBULIN A OXIDATION AT PH 7.5 AFTER PEPSIN DIGESTION EXPERIMENTAL DISCUSSION • In this study, high-resolution mass spectrometry and tandem mass spectrometry were used to confirm the previous NMR data on the conformational changes of β-lactoglobulin A at different pH ranges. • At pH 2, there were only a few oxidized residues, primarily Trp 19 and Tyr 20 located near the N-terminus, and Met 145 and Cys 160 located at the C-terminus. None of the residues in the hydrophobic cavity or the residues in the α-helix were solvent accessible and oxidized. This is consistent with fact that β-lactoglobulin A at this pH is in its monomer and closed conformation. • At pH 4, the protein goes through reversible association forming oligomeric structures. The protein is still in its closed conformation. Therefore the amount of labeling does not change dramatically compared to pH 2. • At pH 6, the protein is in its closed conformation but the E-F loop starts to expose the openingof the cavity. In addition to the oxidation of residues at N and C terminus, one other residue (Ile 71) in the hydrophobic cavity is oxidized. • At pH 8 (Tanford transition), the solvent accessibility and therefore the exposure of residues to oxidation increases dramatically with the opening of the hydrophobic cavity. Apart from residues which are oxidized at the N and C termini, Trp61, Ile 71, Ile 72, and Ile 78 located inside the hydrophobic cavity, and residues in the α-helix (Leu 133, Phe 136, Leu 140) are now oxidized. • In the urea-denatured sample, residues located at different strands are oxidized in addition to all of the oxidized residues seen at different pH ranges, which is consistent with the fact that β-lactoglobulin A isnow in its extended, non-native form. • Stock solutions of 0.5 mg/ml β-lactoglobulin A were prepared in 10 mM NaCl. • Sample solutions were prepared by adjusting the pH of the stock solution with HCl or NaOH to 2, 4, 6 or 8, respectively, in order to obtain different structural states of β-lactoglobulin A. • A urea-denaturation sample was prepared by adding urea to the pH=7.5 sample solution. • Each sample had hydrogen peroxide added (15%), and then was exposed to UV irradiation in a Stratalinker 2400 for 5 minutes to generate hydroxyl radicals (Sharp, Anal. Chem. 2004, 76, 672) • The intact oxidized protein samples were analyzed by ES-FTMS in order to determine the extent of oxidation. • The oxidized samples were digested with either trypsin or pepsin and analyzed by LC-MS/MS (quadrupole ion trap, QIT) in order to determine the site(s) of the oxidized residues. • Turbo SEQUEST was used to determine the putative sites of oxidation, which were verified manually. ESI-FTMS INTRODUCTION • β-lactoglobulin is a major whey protein found in ruminants. In addition to its nutritional function, it also is thought to have a transportation function due to the pH-dependent conformational changes of its structure, as shown below. • During the transport of the hydrophobic molecules in the acidic environment of the stomach, the cavity of β-lactoglobulin A is in its closed conformation. In the basic environment of the intestine, this protein adjusts its conformation to an open position, thereby releasing its cargo. • In this study, a surface mapping technique involving high resolution mass spectrometry is demonstrated for probing the conformational changes of native β-lactoglobulin A in the pH range of 2-8. A urea-denaturated sample of the protein was used as a control to provide the most extended conformation and investigate the most oxidized version of this protein. • The solvent accessibility, and therefore the extent of oxidation, should increase as β-lactoglobulin A adopts to its open conformation in response to the pH increase. Electrospray-Fourier Transform Ion Cyclotron Resonance Mass Spectrometry (9.4 T) • As seen in the above table, the level of oxidation is pH dependent, and increases with pH. • The urea denatured sample was used as a control to observe the most extensive labeling. • There were residues (Met 7, Met 24, Met 107, Met 145,Cys 121 and Cys 160) which were oxidized regardless of the pH. • The residues in the α-helix region were solvent accessible and labeled at pH 8 and urea denaturation, whereas these residues were buried and not oxidized at lower pHs. • The degree of residue labeling appears to correlate with thedifferent structural changes β-lactoglobulin A goes through at different pHs. CONCLUSIONS • Surface mapping is a useful tool for probing pH-dependent conformational changes of β-lactoglobulin A. • Consistent with the published NMR and XRC results, as the pH increases, the number of solvent accessible residues and therefore the oxidation level increases as the hydrophobic cavity opens. • At pH 8, as the E-F loop exposes the hydrophobic cavity, the residues within the cavity were oxidized. • The highest level of oxidation was seen in urea-denatured sample as a control, and the lowest level of oxidation was seen at pH 2 (closed conformation). • In order to better validate and detect more oxidized residues in the peptides which are created, both pepsin and trypsin were used for digestion of this protein. The protease pepsin gave smaller peptides because of its non-specific cleavage. The verification of the spectra with more peptides will be useful in determining the other residues which were not observable with the trypsin digestion. pH-dependent structural states of β-lactoglobulin A β-lactoglobulin A structure N-terminus (310 helical turn)*(residues 1-15) Strand A (residues 16-27) A-B loop (310 helical turn) (residues 28-45) Strand B (residues 46-49) B-C loop (residues 50-52) Strand C (residues 53-63) C-D loop (residues 64-69) Strand D (residues 70-78) D-E loop (residues 79-83) Strand E (residues 84-85) E-F loop (residues 86-89) Strand F (residues 90-98) F-G loop (residues 99-100) Strand G (residues 101-109) G-H loop (310 helical turn) (residues 110-118) Strand H (residues 119-124) H-α helix loop (residues 125-130) α helix (residues 131-142 ) α-strand I (residues 143-150) Strand I (residues 151-157) 310 helical turn (residues 158-162) *Red font indicates regions where oxidized residues were found LC-MS/MS (QIT) Base peak chromatogram of oxidized β-lactoglobulin A at pH 6.65 • pH 2monomer- closed conformation →hydrophobic cavity is closed • pH 3.5(monomer↔dimer equilibrium) • pH > 3.5(shift to dimer conformation) • pH 4 - 5.2(dimer→octamer) • *association into aggregates starts around pH 4.6 (octamer) • Maximal association is at the pH range of 4.40-4.65 • pH 5.2(octamer→dimer) • pH 5.2 - 6 (N-to-Q (Native to Acidic) transition) • pH 6dimer →hydrophobic cavity begins to open • pH > 7(Tanford Transition→ N-to-R (Native to New conformation) • *dimer conformation-open conformation→hydrophobic cavity is completely exposed • pH 10irreversible base-induced unfolding β-lactoglobulin A sequence (Urea-denaturation oxidation coverage) LIVTQTMKGLDIQKVAGTWYSLAMAASDISLLDAQSAPLRVYVEELKPTP EGDLEILLQKWENDECAQKKIIAEKTKIPAVFKIDALNENKVLVLDTDYK KYLLFCMENSAEPEQSLVCQCLVRTPEVDDEALEKFDKALKALPMHIRLS FNPTQLEEQCHI Blue label- Verified residues Red label - Unverified residues Verified residues 4/5 Cysteine 4/4 Methionine 0/2 Histidine 0/4 Tyrosine 1/2 Tryptophan 2/4 Phenylalanine 6/22 Leucine 3/10 Isoleucine 1/8 Proline ACKNOWLEDGEMENTS Oxidized Peptides Identified by Pepsin Digestion Measured Mass Residues Peptide Sequence 943.442 Da 20-28 YSLAMAASD (strand A) 699.704 46-51 LKPTPE (strand B) 756.288 94-99 VLDTDY (strand F) 1233.658 1-11 LIVTQTMKGLD (N-terminus) 1755.052 137-151 KALKALPMHIRLSF (alpha strand I) • Every color-coded residue is a potentially taggable amino acid. • Most of the highly-reactive residues (C, M, aromatics) were found to be oxidized. • The ability to detect and identify the labeled residues depends on the efficiency of the chemical reaction, sample digestion and purification, and the mass spectrometry detection. • DA acknowledge support from the ORNL-UTK Genome Science and Technology Graduate Program • Nathan VerBerkmoes and David L. Tabb are acknowledged for their technical input. • Research was sponsored by the U.S. Department of Energy, Office of Biological and Environmental Research. Oak Ridge National Laboratory is managed by UT-Battelle, LLC for the U.S. Department of Energy under Contract No. DE-AC05-00OR22725.