Download

1 / 28
350 likes | 982 Views
Partioning of the R1 plasmid. A. Structure of the par locus of R1, showung the positions of the parM and parR genes as well as the cis-acting parC site. The transcription start site is in parC site. B. While the plasmid is replicating, ParR bound to parC pairs the

E N D
Partioning of the R1 plasmid A. Structure of the par locus of R1, showung the positions of the parM and parR genes as well as the cis-acting parC site. The transcription start site is in parC site. B. While the plasmid is replicating, ParR bound to parC pairs the replicating plasmids in the cell center and provides a nucleation site for ParM. 1. After replication is complete, the filaments grow by adding successive ATP-ParM subunits to the ends where parC and ParR are bound. ATP is cleaved to ADP to provide energy. 2. After the plasmid copies have been pushed to the ends (poles) of the cell, the ADP-ParM subunits dissociate from one or both ends and the filaments disappear. 3. ParM then recharged with ATP before the plasmids are partitioned again before the next cell division .
Random curing of plasmid with no Par system Each plasmid has an equal chance of going to one of the daughter cells when the cell divides, and occasional cell inherits no plasmids.
II. Conjugation 1. Many plasmids have the ability to transfer themselves and other DNA elements from one cell to another in a process called conjugation. 2. Donor or male strain – the bacterial strain contains the plasmid. In gram-negative bacteria, such cells produce a structure, called the sex pilus, which facilitates conjugation. 3. Recipient or female strain - the bacterial strain that receives the plasmid from a donor strain during conjugation process. 4. Many naturally occurring plasmids can transfer themselves, called self-transmissible plasmids. Self-transmissible plasmids encode all the functions they need to move among cells, and sometimes they can also aid in the transfer of mobilizable plasmids which encode some but not all of the proteins required for transfer. 5. A plasmid will sometimes transfer into a cell that already has a plasmid of the same Inc group, and one of the two plasmids will subsequently be lost.
6. Mechanism of DNA transfer during conjugation in Gram-negative bacteria (1) F+ X F- → F+ + F+ Hfr X F- → Hfr + recombinant (2) Conjugation is a complicated process that requires the products of many genes: (Fig. 5.3) i. tra genes – products required for transfer. In F plasmid, tra genes occupy large region of the plasmid, more than 20 genes. They can be divided into two components: (i) Dtr component – for transfer and replication. They cluster around the oriT. ii. Mpf – for mating-pair formation. They consist of large membrane-associated structure including pilus. (i) Pilus is a tube-like structure that sticks out of the cell surface and constructed of many copies of a single protein called pillin. (ii) The structure of pilus differs markedly among the plasmid transfer system and determine the efficiency of the transfer under various conditions. Ex., long, flexible pilus of F plasmid – liquid; short, thick and rigid – on solid medium.
A simple view of conjugation by a self-transmissible plasmid, the F plasmid
6. Mechanism of DNA transfer during conjugation in Gram-negative bacteria (iii) The channel – Mpf system also encodes a channel or pore through which DNA passes during conjugation. Little is known of its exact structure. (iv) Coupling proteins – The Mpf component is the first to make contact with a recipient cell. Coupling proteins (TraD), part of the Mpf system, provide the specificity for the transport process, so only some of plasmid can be transferred. (v) The relaxosome, consisting of TraY, TraM, TraI and host-encoded IHF bound the the nicked DNA in oriT. (vi) The 5’ end of the nicked strand is bound to a tyrosin in TraI and the 3’ end is associated with TraI in an unspecified way. (vii) TraC, TraD and TraI have ATPase activity.
6. Mechanism of DNA transfer during conjugation in Gram-negative bacteria iii. The central part of the Dtr component of plasmids is the relaxase. (i) The relaxase protein, part of relaxosome nicks DNA at nic site in oriT sequence, is transferred into the recipient cell along with the DNA and reseals the DNA into single-stranded DNA. (ii) Other proteins of relaxosome are not transferred to the recipient cell. (iii) Primase- Primase should not be necessary to prime the replication in donor cell, since the free 3’ hydroxyl end of nicked DNA can be used as primer. But the primase is made in the donor cell, not in the recipient cell. (iv) Why would a plasmid bother to make its own primase and transfer it into the recipient cell if it can use the host cell primase instead? The answer may be that it does this to make itself more promiscuous and able to transfer into a wider variety of bacterial species.
6. Mechanism of DNA transfer during conjugation in Gram-negative bacteria iv. Many naturally occurring plasmids transfer with a high efficiency for only a short time after they are introduced into cells and then transfer only sporadically thereafter. Most of the time the tra genes are repressed, and without the synthesis of pillin and other tra gene function. Plasmids may normally repress their tra genes to prevent infection by male-specific phages or the cells contain plasmid will be killed.
Partial genetic and physical map of 100 kbp F plasmid IS2 and IS3, the insertion sequences; γδ , a transposon (Tn1000); oriV, the origin of replication; oriT, origin of conjugal transfer; tra, the regin encoding numerous tra functions.
The representation of the F transfer apparatus draw from available information
The representation of the F transfer apparatus draw from available information 1. The pilus is assembled with five TraA (pillin) subunits per turn that are inserted into the inner membrane via TraQ and acetylated by TraX. 2. The pilus is extended through a pore constructed of TraB and TraK, a secretion-like protein anchored to the outer membrane by the lipoprotein TraV. 3. TraL seeds the site of pilus assembly and attracts TraC to the pilus base, where it acts to drive assembly in an energy-dependent manner. 4. The two-way arrow indicates the opposing processes of pilus assembly and retraction. 5. The mating-pair formation (Mpf) proteins, include G and N , which aid in mating-pair stabilization (Mps), and S and T, which disrupt mating-pair formation through entry and surface exclusion, respectively. 6. TraF, H, U, W and TrbC, which together with TraN are specific to F-like systems, appear to play a role in pilus retraction, pore formation and mating-pair stabilization.
The representation of the F transfer apparatus draw from available information 7. The relaxosome, consisting of Y, M, l and host- encoded lHF bound to the nicked DNA in oriT is shown interacting with the coupling protein, D, which in turn interacts with B. 8. The 5’ end of the nicked strand is shown bound to a tyrosine (Y) in l, and the 3’ end is shown being associated with l in an unspecified way. 9. The retained, unnicked strand is not shown. 10. TraC, D and l (two sites for both relaxase and helicase) have ATP utilization motifs, represented by curved arrows.
Mechanism of DNA transfer during conjugation, showing the mating-pair formation (Mpf functions) in purple Primase may be encoded either by the host or by the plasmid and injected with the DNA, primes replication of the complementary strand to make the double-stranded circular plasmid DNA in recipient.
Fertility inhibition of the F plasmid (regulation of the tra genes) A. Genetic organization of the tra region. B. The traJ product is a transcriptional activator that is required for transcription of traY-X and finO from promoter PtraY. C. Translation of the traJ mRNA is blocked by hybridization of an antisense RNA, FinP, which is transcribed in the same region from the complementary strand. Protein FinO stabilizes the FinP RNA.
Self-transmissible vs mobilizable plasmids 1. Plasmids that can not transfer themselves but can be transferred by other plasmids are said to be moblizable, and the process by which they are transferred is called mobilization. 2. The simplest mobilizable plasmids merely contain the oriT sequence of a self-transmissible plasmid can be mobilized by that plasmid. 3. The tra genes of the Dtr system of the mobilizable plasmids are called the mob genes and the region required for mobilization is called the mob region. 4. Naturally occurring mobilizable plasmids can often be mobilized by a number of tra systems. 5. Not all mobilizable plasmids can be mobilized by all self- transmissible plasmids, due to the comunnication between the coupling protein of self-transmissible plasmids and relaxase of mobilizable plasmids .
Integration of the F plasmid by homologous recombination between IS2 elements in the plasmid and in the chromosome, forming an Hfr cell
Creation of a prime factor by homologous recombination Prime factor – a plasmid with an incorporated chromosomal gene(s).
Selecting prime factors on the basis of early transfer of a marker A. The hatched region indicates F DNA. B. A prime factor transfers the pro marker early.
Genetic mapping by Hfr crosses • Chromosomal DNA can be transferred by an integrated plasmid. • Once in the recipient cell, the transferred chromosomal DNA may recombine with homologous sequences in the recipient chromosome.
Genetic mapping by Hfr crosses If the trp region is transferred from the donor to the recipient, it can recombine to replace the homologous region in the recipient, giving rise to a Trp+ Arg+ recombinant.
Genetic mapping by Hfr crosses The chromosome is transferred from the donor to the recipient, starting at the position of the integrated self-transmissible plasmid (arrowhead). The plating media used to select the markers are also shown.
Genetic mapping by Hfr crosses 1. Large arrows indicate the known markers used for the Hfr gradient of transfer. 2. The small arrows indicate the position of integration of the plasmid in some Hfr strains, including PK191 (located near the position of hisG at 44 min).
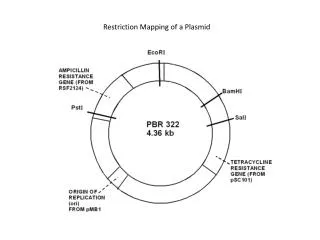
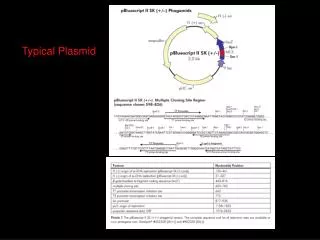
III. Constructing a plasmid cloning vector 1. A cloning vector is an autonomously replicating DNA into which other DNAs can be inserted so that many copies of original piece of DNA can be obtained. 2. Most plasmids, as they are isolated from nature, are too large to be convenient as cloning vectors and/or often do not contain easily selectable genes that can be used to move them from one host to another. 3. A cloning vector can be constructed using the techniques of genetic engineering. A cloning vector should contain three elements: origin of replication (ori), selection marker and unique (multiple) cloning site(s). 4. pBluescript SK as an example.
質體載體(plasmid vector) 質體載體為一個經過修飾過後的質體,將細菌自然存 在的質體移除或加入一些DNA片段而形成。 選殖載體至少必須具備三種不同功能的DNA片段: (1) 複製的起始點(origin of DNA replication): 為質體開始複製的地方。 (2) 多選殖位點(multiple cloning sites,MCS): 為一小段DNA,內含有多個不同的限制酶的切 位,以供外源DNA片段插入。 (3) 篩選標誌(selection marker)::以 pBluescript SK 載體例,其篩選標誌為合成分解氨比黴素(ampicilin) 的基因。當大腸桿菌含有此載體時,可以在含有氨比黴 素的環境下生長。lacZ’基因為另一種常用的篩選 標誌,可用來確定是否有一段DNA插入載體之MCS內。
The rationale behind insertional inactivation of the lacZ’ gene carried by pUC8 (a) The bacterial and plasmid genes complement each other to produce a functional β–galactosidase molecule. (b) Recombinants are screened by plating onto agar containing X-gal and IPTG.
Plasmid amplification The copy number of some plasmid vectors (for example, the pUC8 or pBluescript SK ) can be increased by following procedures: 1. Grow the plasmid-containing bacteria to early log phase. 2. Adding the antibiotic chlorophenicol (CM) 3. Culturing the bacteria over night. Why?