Download
1 / 70
700 likes | 908 Views
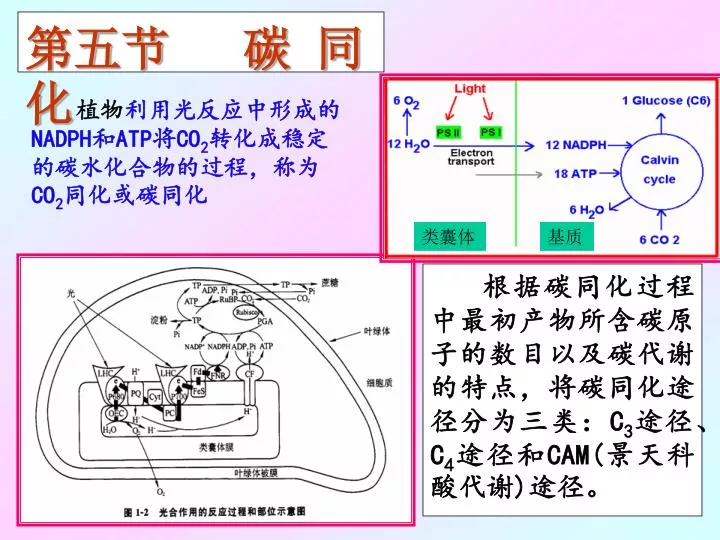
第五节 碳 同 化. 植物 利用光反应中形成的 NADPH 和 ATP 将 CO 2 转化成稳定的碳水化合物的过程,称为 CO 2 同化或碳同化. 类囊体. 基质. 根据碳同化过程中最初产物所含碳原子的数目以及碳代谢的特点,将碳同化途径分为三类: C 3 途径、 C 4 途径和 CAM( 景天科酸代谢 ) 途径。. 一 .C 3 途径. 1946 年,美国加州大学放射化学实验室的卡尔文 (M.Calvin) 和本森 (A.Benson) 等人采用了两项新技术:

E N D
第五节 碳 同 化 植物利用光反应中形成的NADPH和ATP将CO2转化成稳定的碳水化合物的过程,称为CO2同化或碳同化 类囊体 基质 根据碳同化过程中最初产物所含碳原子的数目以及碳代谢的特点,将碳同化途径分为三类:C3途径、C4途径和CAM(景天科酸代谢)途径。
一.C3途径 1946年,美国加州大学放射化学实验室的卡尔文(M.Calvin)和本森(A.Benson)等人采用了两项新技术: • (1)14C同位素标记与测定技术(可排除原先存在于细胞里的物质干扰,凡被14C标记的物质都是处理后产生的); • (2)双向纸层析技术(能把光合产物分开)。 • 选用小球藻等单细胞的藻类作材料,藻类不仅在生化性质上与高等植物类似,且易于在均一条件下培养,还可在试验所要求的时间内快速地杀死。
试验分以下几步进行: (1)饲喂14CO2与定时取样 向正在进行光合作用的藻液中注入14CO2使藻类与14CO2接触,每隔一定时间取样,并立即杀死。 H14CO3-+H+→14CO2+H2O 图用来研究光合藻类CO2固定仪器的图解
(2)浓缩样品与层析 用甲醇将标记化合物提取出来,将样品浓缩后点样于层析纸上,进行双向纸层析,使光合产物分开 (3)鉴定分离物 采用放射自显影技术,鉴定被14CO2标记的产物并测定其相对数量。 (4)设计循环图 根据被14C标记的化合物出现时间的先后,推测生化过程。根据图D所显示的结果,即短时间内(5秒,最终到0.5秒钟)14C标记物首先出现在3-磷酸甘油酸(PGA)上,说明PGA是光合作用的最初产物。 用纸层析和放射自显影技术追踪被14CO2标记的产物
起先猜测CO2是与某一个2碳的片断结合生成3碳的PGA,然而情况并非如此。 当光下把CO2浓度突然降低,作为CO2受体的化合物会积累起来。这一化合物被发现是含有5个C的核酮糖-1,5-二磷酸(RuBP),当它接受CO2后,分解为2个PGA分子。 光合试验中RuBP与PGA相互转化 经过10多年周密的研究,卡尔文等人终于探明了光合作用中从CO2到蔗糖的一系列反应步骤,推导出一个光合碳同化的循环途径,这条途径被称为卡尔文循环或Calvin-Benson循环 。 由于这条途径中CO2固定后形成的最初产物PGA为三碳化合物,所以也叫做C3途径或C3光合碳还原循环,并把只具有C3途径的植物称为C3植物。 此项研究的主持人卡尔文获得了1961年诺贝尔化学奖。
光合碳还原循环 • 代谢产物名:RuBP.核酮糖1,5二磷酸; PGA.3-磷酸甘油酸; BPGA.1,3二磷酸甘油酸; GAP.甘油醛-3-磷酸; DHAP.二羟丙酮磷酸; FBP.果糖-1,6-二磷酸; F6P.果糖-6-磷酸; E4P.赤藓糖-4-磷酸; SBP.景天庚酮糖-1,7-二磷酸; S7P.景天庚酮糖-7-磷酸; R5P.核糖-5-磷酸; Xu5P.木酮糖-5-磷酸; Ru5P.核酮糖-5-磷酸; G6P.葡萄糖-6-磷酸; TPP.硫胺焦磷酸; TPP-C2.TPP羟基乙醛 参与反应的酶:(1)核酮糖二磷酸羧化酶/加氧酶(Rubisco); (2)3-磷酸甘油酸激酶(PGAK); (3)NADP-甘油醛-3-磷酸脱氢酶; (4)丙糖磷酸异构酶; (5)(8)醛缩酶; (6)果糖-1,6-二磷酸(酯)酶(FBPase); (7)(10)(12)转酮酶; (9)景天庚酮糖1,7-二磷酸(酯)酶(SBPase); (11)核酮糖-5-磷酸表异构酶; (13)核糖-5-磷酸异构酶; (14)核酮糖-5-磷酸激酶(Ru5PK)
(一) C3途径的反应过程 • C3途径是光合碳代谢中最基本的循环,是所有放氧光合生物所共有的同化CO2的途径。 1.过程 • 整个循环如图所示,由RuBP开始至RuBP再生结束,共有14步反应,均在叶绿体的基质中进行。 • 全过程分为羧化、还原、再生3个阶段。 一分子C02固定需要消耗2分子NADPH和3分子ATP
羧化阶段 指进入叶绿体的CO2与受体RuBP结合,并水解产生PGA的反应过程。 以固定3分子CO2为例: 3RuBP+3CO2+3H2O Rubisco 6PGA + 6H+ 核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)具有双重功能,既能使RuBP与CO2起羧化反应,推动C3碳循环,又能使RuBP与O2起加氧反应而引起C2氧化循环即光呼吸。 羧化阶段分两步进行,即羧化和水解: 在Rubisco作用下RuBP的C-2位置上发生羧化反应形成2-羧基-3-酮基阿拉伯糖醇-1,5-二磷酸,它是一种与酶结合不稳定的中间产物,被水解后产生2分子PGA。
(2)还原阶段 • 指利用同化力将3-磷酸甘油酸还原为甘油醛-3-磷酸的反应过程 • 6PGA+6ATP+6NADPH+ 6H+→→6GAP+6ADP+6NADP+ + 6Pi • 有两步反应:磷酸化和还原。磷酸化反应由3-磷酸甘油酸激酶催化 • 羧化反应产生的PGA是一种有机酸,要达到糖的能级,必须使用光反应中生成的同化力,ATP与NADPH能使PGA的羧基转变成GAP的醛基。当CO2被还原为GAP时,光合作用的贮能过程便基本完成。
(3)再生阶段 • 指由甘油醛-3-磷酸重新形成核酮糖-1,-5-二磷酸的过程5GAP+3ATP+2H2O→→→3RuBP+3ADP+2Pi+3H+ • 这里包括形成磷酸化的3,4,5,6和7碳糖的一系列反应。最后由核酮糖-5-磷酸激酶(Ru5PK)催化,消耗1分子ATP,再形成RuBP。
光合碳还原循环 羧化 还原 再生
C3途径的总反应式 3CO2+5H2O+9ATP+6NADPH→GAP+9ADP+8Pi+6NADP++3H+ • 可见,每同化一个CO2需要消耗3个ATP和2个NADPH,还原3个CO2可输出1个磷酸丙糖(GAP或DHAP) • 固定6个CO2可形成1个磷酸己糖(G6P或F6P)。 • 形成的磷酸丙糖可运出叶绿体,在细胞质中合成蔗糖或参与其它反应; • 形成的磷酸己糖则留在叶绿体中转化成淀粉而被临时贮藏。 再
二、C4 途 径 (一) C4 途径的发现 • 自20世纪50年代卡尔文等人阐明C3途径以来,曾认为不管是藻类还是高等植物,其CO2固定与还原都是按C3途径进行的。 • 1954年,哈奇(M.D.Hatch)等人用甘蔗叶实验,发现甘蔗叶片中有与C3途径不同的光合最初产物,但未受到应有的重视。 • 1965年,美国夏威夷甘蔗栽培研究所的科思谢克(H.P.Kortschak)等人报道,甘蔗叶中14C标记物首先出现于C4二羧酸,以后才出现在PGA和其他C3途径中间产物上,而且玉米、甘蔗有很高的光合速率,这时才引起人们广泛的注意。 • 1966-1970年,澳大利亚的哈奇和斯莱克(C.R.Slack) 重复上述实验,进一步地追踪14C去向,探明了14C固定产物的分配以及参与反应的各种酶类,于70年代初提出了C4-双羧酸途径,简称C4途径,也称C4光合碳同化循环,或叫Hatch-Slack途径。 • 至今已知道,被子植物中有20多个科约近2000种植物按C4途径固定CO2,这些植物被称为C4植物(C4 plant)。
C4植物 高梁 玉米 粟(millet)的穗形, “谷子”,去皮后称“小米” 甘蔗田 苋菜
(二)C4植物叶片结构特点 • 栅栏组织与海绵组织分化不明显,叶片两侧颜色差异小。 • 有两类光合细胞:叶肉细胞和维管束鞘细胞(BSC)。 • C4植物维管束分布密集,间距小,每条维管束都被发育良好的大型BSC包围,外面又密接1-2层叶肉细胞,这种呈同心圆排列的BSC与周围的叶肉细胞层被称为“花环”(Kranz,德语)结构,C4植物的BSC中含有大而多的叶绿体,线粒体和其它细胞器也较丰富。 • BSC与相邻叶肉细胞间的壁较厚,壁中纹孔多,胞间连丝丰富。这些结构特点有利于MC与BSC间的物质交换,以及光合产物向维管束的就近转运。 • 两类光合细胞中含有不同的酶类,叶肉细胞中含有PEPC以及与C4二羧酸生成有关的酶;而BSC中含有Rubisco等参与C3途径的酶、乙醇酸氧化酶以及脱羧酶。在这两类细胞中进行不同的生化反应。
C4植物玉米花环状维管束细胞的解剖结构图。紧密的维管束鞘四周被大的维管束鞘细胞环绕包围。在这类作物中大的叶绿体分布在维管束鞘细胞的外围,维管束细胞被叶肉细胞包围。C4植物玉米花环状维管束细胞的解剖结构图。紧密的维管束鞘四周被大的维管束鞘细胞环绕包围。在这类作物中大的叶绿体分布在维管束鞘细胞的外围,维管束细胞被叶肉细胞包围。
(三)C4途径的反应过程 C4途径中的反应虽因植物种类不同而有差异,但基本上可分为羧化、还原或转氨、脱羧和底物再生四个阶段(图22,C)。 • ①羧化反应 在叶肉细胞中磷酸烯醇式丙酮酸(PEP)与HCO-3在PEPC催化下形成草酰乙酸(OAA); • ②还原或转氨作用OAA被还原为苹果酸(Mal),或经转氨作用 形成天冬氨酸(Asp); • ③脱羧反应C4酸通过胞间连丝移动到BSC,在BSC中释放CO2,CO2由C3 途径同化; • ④底物再生 脱羧形成的C3酸从BSC运回叶肉细胞并再生出CO2受体PEP。 图22 C4植物叶的结构以及 C4 植物光合碳代谢的基本反应 ( )内为酶名; PEPC.PEP羧化酶; PPDK.丙酮酸磷酸二激酶
1.羧化阶段 • 由PEPC催化叶肉细胞中的磷酸烯醇式丙酮酸(PEP)与HCO3-羧化,形成OAA。 • 空气中的CO2进入叶肉细胞后先由碳酸酐酶 (CA)转化为HCO3-, HCO3- 被PEP固定在OAA的C4羧基上 • CO2 +H2O CA HCO3-+ H+ • PEPC是胞质酶,主要分布在叶肉细胞的细胞质中,分子量400 000,由四个相同亚基组成。PEPC无加氧酶活性,因而羧化反应不被氧抑制。
2.还原或转氨阶段 • OAA被还原成苹果酸或经转氨作用形成天冬氨酸。 • (1)还原反应 由NADP-苹果酸脱氢酶催化,将OAA还原为Mal,该反应在叶肉细胞的叶绿体中进行: • 苹果酸脱氢酶为光调节酶,可通过Fd-Td系统调节其活性。 • (2)转氨作用 由天冬氨酸转氨酶催化,OAA接受谷氨酸的NH2基,形成天冬氨酸,该反应在细胞质中进行。
3.脱羧阶段 根据植物所形成的初期C4二羧酸的种类以及脱羧反应参与的酶类,可把C4途径分为三种亚类型: • ①依赖NADP的苹果酸酶的苹果酸型(NADP-ME型); • ②依赖NAD的苹果酸酶的天冬氨酸型(NAD-ME型); • ③具有PEP羧激酶的天冬氨酸型(PCK型)。 NADP-ME型初期产物为Mal,而NAD-ME型与PCK型初期产物为Asp。 叶绿体 线粒体 细胞质 三种亚类型叶绿体的结构及其在BSC中的排列有所不同。就禾本科植物而言,NAD-ME型植物,叶绿体在BSC中向心排列,而NADP-ME与PCK型,叶绿体在BSC中离心排列;另外NADP-ME型BSC中叶绿体的基粒不发达,PSⅡ活性低。
4.底物再生阶段 • C4二羧酸脱羧后形成的Pyr运回叶肉细胞,由叶绿体中的丙酮酸磷酸二激酶(PPDK)催化,重新形成CO2受体PEP。 • NAD-ME型和PCK型形成的丙氨酸在叶肉细胞中先转为丙酮酸,然后再生成PEP。 • 此步反应要消耗2个ATP(因AMP变成ADP再要消耗1个ATP)。 • PPDK在体内存在钝化与活化两种状态,它易被光活化,光下该酶的活性比暗中高20倍。 • 由于PEP底物再生要消耗2个ATP,这使得C4植物同化1个CO2需消耗5个ATP与2个NADPH。
(四) C4途径的意义 • 在高温、强光、干旱和低CO2条件下,C4植物显示出高的光合效率。 C4植物具较高光合速率的因素有: • C4植物的叶肉细胞中的PEPC对底物HCO-3的亲和力极高,细胞中的HCO3-浓度一般不成为PEPC固定CO2的限制因素; • C4植物由于有“CO2泵”浓缩CO2的机制,使得BSC中有高浓度的CO2,从而促进Rubisco的羧化反应,降低了光呼吸,且光呼吸释放的CO2又易被再固定; • 高光强又可推动电子传递与光合磷酸化,产生更多的同化力,以满足C4植物PCA循环对ATP的额外需求; • 鞘细胞中的光合产物可就近运入维管束,从而避免了光合产物累积对光合作用可能产生的抑制作用。 • 但是C4植物同化CO2消耗的能量比C3植物多,也可以说这个“CO2泵”是要由ATP来开动的,故在光强及温度较低的情况下,其光合效率还低于C3植物。可见C4途径是植物光合碳同化对热带环境的一种适应方式。
三、景天科酸代谢途径 (一)CAM在植物界的分布与特征 景天科等植物有一个很特殊的CO2同化方式:夜间固定CO2产生有机酸,白天有机酸脱羧释放CO2,用于光合作用,这样的与有机酸合成日变化有关的光合碳代谢途径称为CAM(Crassulacean acid metabolism)途径 景天科酸代谢途径 (1)PEP羧化酶(2)苹果酸脱氢酶(3)NADP+— 苹果酸酶
CAM最早是在景天科植物中发现的,目前已知在近30个科,1万多个种的植物中有CAM途径,主要分布在景天科、仙人掌科、兰科、凤梨科、大戟科、番杏科、百合科、石蒜科等植物中。其中凤梨科植物达1千种以上,兰科植物达数千种,此外还有一些裸子植物和蕨类植物。 • CAM植物起源于热带,往往分布于干旱的环境中,多为肉质植物,具有大的薄壁细胞,内有叶绿体和液泡,然而肉质植物不一定都是CAM植物。 • 常见的CAM植物有菠萝、剑麻、兰花、百合、仙人掌、芦荟、瓦松等。
剑麻 龙舌兰 落地生根 绯牡丹 芦荟 昙花
CAM植物-瓦松属 瓦松属1
多肉质植物 鸡冠掌 静夜 红司 锦晃星
(二) CAM代谢的反应过程 • 典型CAM植物一天中的CO2固定速率,气孔导度、苹果酸和葡聚糖(如淀粉)含量变化以及代谢过程如图24和图25所示。 • 图24 CAM植物气孔导度、CO2固定、苹果酸和葡聚糖含量的日变化 • 阶段Ⅰ.夜间CO2吸收旺盛期; • 阶段Ⅱ.日间开始CO2吸收期; • 阶段Ⅲ.日间CO2交换停止期; • 阶段Ⅳ.日间结束CO2吸收期。
CO2固定昼夜可变化分为4个阶段: 1.阶段Ⅰ PEPC的羧化阶段。夜间气孔开启,CO2被PEPC固定生成草酰乙酸,后者还原成苹果酸贮存于液泡。此间CO2固定约占全天固定CO2的3/4左右,而白天贮存的葡聚糖在此期间用于形成PEP。 2.阶段Ⅱ由PEPC羧化转向Rubisco羧化的阶段。白昼开始时气孔导度与CO2吸收出现一个高峰,此期间C4途径与C3途径同时进行,苹果酸积累中止。 3.阶段Ⅲ Rubisco同化CO2阶段。日间气孔关闭,停止从外界吸收CO2,苹果酸从液泡转移至细胞质,氧化脱羧。脱羧释放的CO2进入叶绿体被 C3途径同化。 4.阶段Ⅳ由Rubisco羧化转向由PEPC羧化的阶段。苹果酸脱羧降至最低点,气孔开始张开,CO2吸收增加,且由C3途径过渡至C4途径,从而又重复下一个昼夜变化周期。
图8.12 景天科酸代谢途径(CAM)。光合反应吸收的CO2暂时的分离:夜间CO2的吸收和固定,白天内部释放的CO2进行脱酸和再固定。
(1)夜间固定CO2,产生苹果酸,贮藏于液泡中。 (2)白天有机酸脱羧,参加卡尔文循环。
上述这些阶段的划分不是截然分明的,而是逐渐地由一个阶段过渡到下一个阶段。上述这些阶段的划分不是截然分明的,而是逐渐地由一个阶段过渡到下一个阶段。 • CAM植物与C4植物固定与还原CO2的途径基本相同,二者的差别在于: • C4植物是在同一时间(白天)和不同的空间(叶肉细胞和维管束鞘细胞)完成CO2固定(C4途径)和还原(C3途径)两个过程; • 而CAM植物则是在不同时间(黑夜和白天)和同一空间(叶肉细胞)完成上述两个过程的。 • CAM植物由于白天气孔关闭、苹果酸脱羧、细胞间的CO2/O2比例高、以及CO2再固定率高,因而表观光呼吸较低。
四、C3、C4、CAM植物的特性比较 (一)C3、C4、CAM植物的特性比较 C3植物,C4植物和CAM植物的光合作用与生理生态特性有较大的差异。见表4-4。
(二)C3、C4、CAM植物的相互关系 • 从生物进化的观点看,C4植物和CAM植物是从C3植物进化而来的。 • 在陆生植物出现的初期,大气中CO2浓度较高,O2较少,光呼吸受到抑制,故C3途径能有效地发挥作用。 • 随着植物群体的增加,O2浓度逐渐增高,CO2浓度逐渐降低,一些长期生长在高温、干燥气候下的植物受生态环境的影响,也逐渐发生了相应的变化。如出现了花环结构,叶肉细胞中的PEPC和磷酸丙酮酸二激酶含量逐步增多,形成了有浓缩CO2机制的C4-二羧酸循环,形成了C3-C4中间型植物乃至C4植物,或者形成了白天气孔关闭,抑制蒸腾作用,晚上气孔开启,吸收CO2的CAM植物。 • 不过,不论是哪一种光合碳同化类型的植物,都具有C3途径,这是光合碳代谢的基本途径。C4途径、CAM途径以及光呼吸途径只是对C3途径的补充。 • 由于长期受环境的影响,使得在同一科属内甚至在同一植物中可以具有不同的光合碳同化途径。
例如禾本科黍属的56个种内有C4植物种47个,C3植物种8个, C3-C4中间类型1个; • 在大戟属和碱蓬属内,则同时存在C3、C4和CAM植物。 • 禾本科的毛颖草在低温多雨地区为C3植物,而在高温少雨地区为C4植物。 • C3植物感病时往往会出现C4植物的特征,如C3植物烟草感染花叶病毒后,PEPC代替了被抑制了的Rubisco,在幼叶中出现了C4途径。 • 玉米幼苗叶片具有C3特征,至第五叶才具有完全的C4特性。 • C4植物衰老时,会出现C3植物的特征。 • 也有一些肉质植物在水分胁迫条件下由C4途径转变为CAM途径。CAM植物则有专性和兼性之分。 • 总之,不同碳代谢类型之间的划分不是绝对的,它们在一定条件下可互相转化,这也反映了植物光合碳代谢途径的多样性、复杂性以及在进化过程中植物表现出的对生态环境的适应性。
(三) 光合碳代谢多样性意义 (1)植物为了适应不同的环境条件,植物的起源不同,进化出不同的光合碳代谢类型。 (2)同一植物在不同的环境条件下,不同植物在不同的环境条件下。 禾本科植物毛颖草在低温多雨地区为C3植物,而在高温干旱地区为C4植物。 (3)同一植物在不同的生理状况下,不同的生育时期。C3植物在感病时各自为往往会出现C4植物的特征。烟草感染花叶病毒后,PEPC代替了被抑制了的Rubisco,在幼叶中出现了C4途径。玉米幼苗叶片具有C3特征,到第5叶才具有完全的C4特征。C4植物衰老时会出现C3植物的特征。CAM植物则有很好的随环境变化 而适应的特征。
第六节 光 呼 吸 光呼吸:植物的绿色细胞在光照下有吸收氧气,释放CO2的反应。 (一)光呼吸的发现 1920年瓦伯格在用小球藻做实验时发现,O2对光合作用有抑制作用,这种现象被称为瓦伯格效应(War burg effect)。这实际上是氧促进光呼吸的缘故。
1955年德克尔(J.P.Decher)用红外线CO2气体分析仪测定烟草光合速率时,观察到对正在进行光合作用的叶片突然停止光照,断光后叶片有一个CO2快速释放(猝发)过程。 CO2猝发(CO2 outburst)现象实际上是光呼吸的“余辉”,即在光照下所形成的光呼吸底物尚未立即用完,在断光后光呼吸底物的继续氧化。 现在通常把1955年作为发现光呼吸的年代。 • 1971年托尔伯特(Tolbert)阐明了光呼吸的代谢途径。
(二)光呼吸的生化途径 现在认为光呼吸的生化途径是乙醇酸的代谢,主要证据: 以上两步反应在叶绿体中进行
乙醇酸从叶绿体转入过氧化体,由乙醇酸氧化酶催化氧化成乙醛酸, • 乙醛酸经转氨作用转变为甘氨酸,甘氨酸在进入线粒体后发生氧化脱羧和羟甲基转移反应转变为丝氨酸,丝氨酸再转回过氧化体,并发生转氨作用,转变为羟基丙酮酸,后者还原为甘油酸,转入叶绿体后,在甘油酸激酶催化下生成的3-磷酸甘油酸又进入C3途径,整个过程构成一个循环。 • 其中耗氧反应部位有两处,一是叶绿体中的Rubisco加氧反应,二是过氧化体中的乙醇酸氧化反应。 • 脱羧反应则在线粒体中进行,2个甘氨酸形成1个丝氨酸时脱下1分子CO2。 • 图光呼吸途径及其在细胞内的定位 • ①Rubisco; ②磷酸乙醇酸磷酸(酯)酶; ③乙醇酸氧化酶; ④谷氨酸-乙醛酸转氨酶; ⑤丝氨酸-乙醛酸氨基转移酶; ⑥甘氨酸脱羧酶⑦丝氨酸羟甲基转移酶 ⑧羟基丙酮酸还原酶; ⑨甘油酸激酶
图光呼吸途径及其在细胞内的定位 • ①Rubisco; ②磷酸乙醇酸磷酸(酯)酶; ③乙醇酸氧化酶; ④谷氨酸-乙醛酸转氨酶; ⑤丝氨酸-乙醛酸氨基转移酶; ⑥甘氨酸脱羧酶⑦丝氨酸羟甲基转移酶 ⑧羟基丙酮酸还原酶; ⑨甘油酸激酶
从RuBP到PGA的整个反应总方程式为: RuBP+15O2+11H2O+34ATP+15NADPH+10Fdred 5CO2+34ADP+36Pi+15NADP++10Fdox+9H+ 因为光呼吸底物乙醇酸和其氧化产物乙醛酸,以及后者经转氨作用形成的甘氨酸皆为C2化合物,因此光呼吸途径又称为C2光呼吸碳氧化循环,简称C2循环。
