Download
1 / 11
110 likes | 316 Views
INTERMEDIATES IN THE REACTION CYCLE OF CYTOCHROME P450cam. V. Schünemann, Technische Universität Kaiserslautern, Germany C. Jung, KKS Ultraschall AG, Switzerland A.X. Trautwein, Universität zu Lübeck, Germany. Motivation

E N D
INTERMEDIATES IN THE REACTION CYCLE OF CYTOCHROME P450cam V. Schünemann, Technische Universität Kaiserslautern, Germany C. Jung, KKS Ultraschall AG, Switzerland A.X. Trautwein, Universität zu Lübeck, Germany
Motivation The physiologically important enzyme superfamily cytochrome P450 catalyzes a variety of reactions, such as aliphatic and aromatic hydroxylations, epoxidations, heteroatom oxidation, and N- and O-dealkylation, by transfer of an active oxygen atom from its heme unit to the substrates. All enzymes of the cytochrome P450 family have a hydrophobic binding pocket, in which resides a protoporphyrine IX with its iron center being coordinated to an axial cysteine ligand [1]. The enzyme cytochrome P450cam from Pseudomonas putida hydroxylases (1R)-camphor as natural substrate and is regarded to be a representative enzyme for the whole P450 family. Mössbauer spectroscopic studies on this enzyme have been performed by Debrunner and coworkers in the seventies [2] of the last century. The results of these studies provided insight into the catalytic mechanism of the enzyme even before the crystal structure of the resting state of P450cam was published [3]. HO Fig. 1 Cytochrome P450cam (left) adds an oxygen atom O at the 5-exo position of camphor (C10H16O, right).
The cytochrome P450 reaction cycle The postulated enzymatic reaction cycle is shown in Fig. 2. In the resting state of the enzyme the catalytically active heme iron center acquires the ferric low-spin state (S=1/2). After binding of the substrate camphor to the amino acid residue Tyr96 inside the heme pocket, the iron changes from the ferric low-spin to the ferric high-spin state (S=5/2). The transfer of the first electron originating from NAD(P)H via redox proteins (flavin and iron-sulfur proteins) reduces the iron to the ferrous high-spin state (S=2). Subsequent binding of molecular oxygen to the iron forms a diamagnetic FeO2 center, similar to the oxygenated state of myoglobin. The transfer of a second electron initiates catalytic steps which lead to an iron-oxo intermediate called compound I (cpd I). It is this intermediate which inserts the active oxygen atom into the substrate camphor. Fig. 2 The reaction cycle of cytochrome P450. The “shunt“ reaction used in this study is depicted in red.
S Fe(IV)=1 S Radical=1/2 The nature of the intermediate cpd I Neither the electronic structure of cpd I of P450cam nor of any other P450 has been unambiguously determined up to now, but it is generally assumed that the putative cpd I in cytochrome P450 should contain the same electron and spin distribution as is observed for cpd I of peroxidases [4], catalases and many synthetic cpd I analogues [5]. In these systems one oxidation equivalent resides on the Fe(IV)=O unit (d4, S=1) and one is located on the porphyrin (S'=1/2), constituting a magnetically coupled ferryl iron-oxo porphyrin -cation radical system. For other enzymes like cytochrome c peroxidase a ferryl Fe(IV) coupled to an organic radical on the protein (SRadical=1/2) has been proposed [6]. R-H + O2+ 2e- + 2H+ R-OH + H2O Fig. 3 The Intermediate cpd I (left) inserts oxygen into the substrate R-H via the reaction shown on the right.
Addition of peroxy acetic acid to native Cyt P450cam • Investigation with • Mössbauer Spectroscopy • Multi-Frequency EPR Spectroscopy Experimental techniques Reaction intermediates can be trapped by rapid freeze-quench experiments (Fig. 4) . This technique allows to prepare samples for Mössbauer- and electron paramagnetic resonance (EPR-) spectroscopy. In order to detect cpd I in cytochrome P450cam we have used the “shunt” reaction (see Fig. 2) with peroxy acetic acid. • Freezing of the reaction mixture within milliseconds Fig. 4
The start: Mössbauer Spectroscopy on camphor-free cytochrome P450cam : The Mössbauer spectrum obtained at 4.2K and B=20 mT of cytochrome P450cam without the substrate camphor is shown in Fig. 5. The observed magnetic splitting and the corresponding EPR spectrum (Fig. 6) are consistent with a ferric low-spin state of the iron. The Mössbauer spectrum has been analyzed by the spin-Hamiltonian formalism (solid line in Fig. 5) and the parameters are shown in Tab. 1 [7]. Fig. 6 Fig. 5 Table 1. Mössbauer parameters used for the spin Hamiltonian simulations of the ferric low-spin species of cytochrome P450cam (solid lines in Figs. 5 and 7).
Freeze-quench experiments I: 57Fe(III)P450cam + substrate camphor + peroxy acetic acid • The Mössbauer spectrum obtained at 4.2K and B=20 mT of cytochrome P450cam with the substrate camphor after reaction with peroxy acetic acid for 8 ms is shown in Fig. 7. The spectrum displays an S=5/2-1/2 equilibrium which is characteristic for the substrate bound P450cam, but no Fe(IV) has been detected. The corresponding EPR spectrum (Fig. 8) also shows an S=5/2-1/2 equilibrium. No characteristic signals for cpd I with S=1/2 or S=3/2 have been observed, only very small amounts of an organic radical (0.6%). Obviously the addition of peroxy acetic acid does not significantly affect the Mössbauer- and EPR pattern of the substrate bound cytochrome P450cam. Fig. 8 Fig. 7 Table 2. Mössbauer parameters used for the spin Hamiltonian simulation of the ferric –high-spin species of cytochrome P450cam shown in Fig. 7 (taken from [8]).
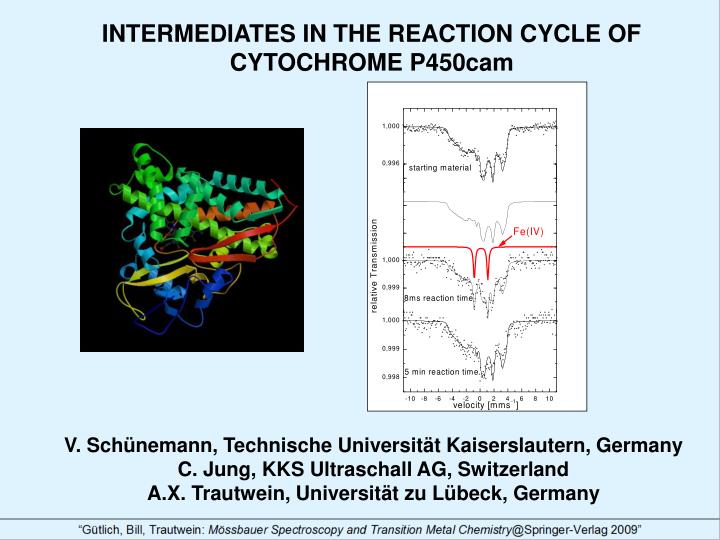
Freeze-quench experiments II: Substrate-free 57Fe(III)P450cam + peroxy acetic acid The Mössbauer spectrum obtained at 4.2K and B=20 mT of substrate-free P450cam after reaction with peroxy acetic acid for 8 ms is shown in Fig. 9. In addition to the ferric-low spin form (black solid line with parameters given in Table 1) the spectrum displays a doublet (red line, 132%) with =0.13 mms-1 and EQ=1.94 mms-1 characteristic for a ferryl species ((Fe(IV)=O)2+ ; S=1) [9] which has been shown to be protonated [10]. The pattern of the starting material is restored after 5 min reaction time. This shows that the ferryl species is indeed a transient intermediate. Fig. 9 The corresponding EPR spectrum (Fig. 10) shows an intense radical signal with a spectroscopic signature pointing towards a tyrosyl radical. Fig. 10 Tyrosyl radical?
2- Fig. 11 Cytochrome P450cam substrate-bound Assignment of the radical signal to Tyr96 A multi-frequency EPR study of the freeze-quenched intermediate obtained from reaction of substrate-free cytochrome P450cam and its Y96F and Y96F-Y75F mutants with peroxy acids has been performed [11]. High-field EPR studies at 285 and 94 GHz on freeze-quenched wild type and Y96F samples reveal g-tensor components for the radical (stretched gx-values from 2.0078-2.0064, gy = 2.0043, and gz = 2.0022), which are fingerprints for tyrosine radicals. From the simulation of the hyperfine structure in the 94 GHz EPR spectra the signals have been assigned to Y96 in the wild type and to Y75 in the Y96F mutant. We suggest that a transiently formed Fe(IV)=O porphyrin--cation radical intermediate in P450cam is reduced by intramolecular electron transfer from these tyrosines within 8 ms (Fig. 11). Such an intramolecular electron transfer may be a general phenomenon in cpd I formation (Fig.12). Fig. 12
Summary A complementary Mössbauer- and EPR-study of the time dependence of the reaction of substrate free P450cam with peracetic acid within a time region ranging from 8 ms up to 5 min has been presented. An Fe(IV) species as well as a tyrosyl radical residing on the amino acid residue Tyr96 have been identified as reaction intermediates. These species possibly are formed by the reduction of compound I by means of transferring an electron from Tyr 96 to the heme moiety. Readers who are interested in the field of reaction intermediates of cytochrome P450 are referred to a review article by Jung et al. [12].
References [1] D.F.V. Lewis, Cytochromes P450- Structure, Function and Mechanism, Taylor & Francis Ltd, London, 1996. [2] M. Sharrock, P.G. Debrunner, C. Schulz, J.D. Lipscomb, V. Marshall and I.C.Gunsalus, Biochim. Biophys. Acta 420, 8 (1976) [3] T.L. Poulos, B.C. Finzel and A.J. Howard, Biochemistry 25, 5314 (1986). [4] R. Rutter, L.P. Hager, H. Dhonau, M. Hendrich, M. Valentine and P. Debrunner, Biochemistry 23, 6809 (1984) [5] E. Bill, Hyp. Int. 90, 143 (1994) [6] G. Lang, K. Spartalian and T. Yonetani, Biochim. Biophys. Acta 451, 250 (1976) [7] M. Sharrock, P.G. Debrunner, C. Schulz, J.D. Lipscomb, V. Marshall and I.C. Gunsalus Biochim. Biophys. Acta 420, 8 (1976) [8] V. Schünemann, C. Jung, J. Terner, A.X. Trautwein and R. Weiss, J. Inorg. Biochem. 91, 596 (2002) [9] V. Schünemann, C. Jung, A.X. Trautwein, D. Mandon and R. Weiss, FEBS Lett. 479, 149 (2000) [10] R.K. Behan, L.M. Hoffart, K.L. Stone, C. Krebs and M.T. Green, J. Am. Chem. Soc. 128(35), 11471 (2006) [11] V. Schünemann, F. Lendzian, C. Jung, J. Contzen, A.-L. Barra, S.G. Sligar and A.X. Trautwein, J. Biol. Chem. 279 No.12, 10919 (2004) [12] C. Jung, V. Schünemann, F. Lendzian, A. X. Trautwein, J. Contzen, M. Galander, L. H. Böttger, M. Richter and A.-L. Barra, Biol. Chem. 386 No. 10, 1043 (2005)