Download
1 / 37
450 likes | 843 Views
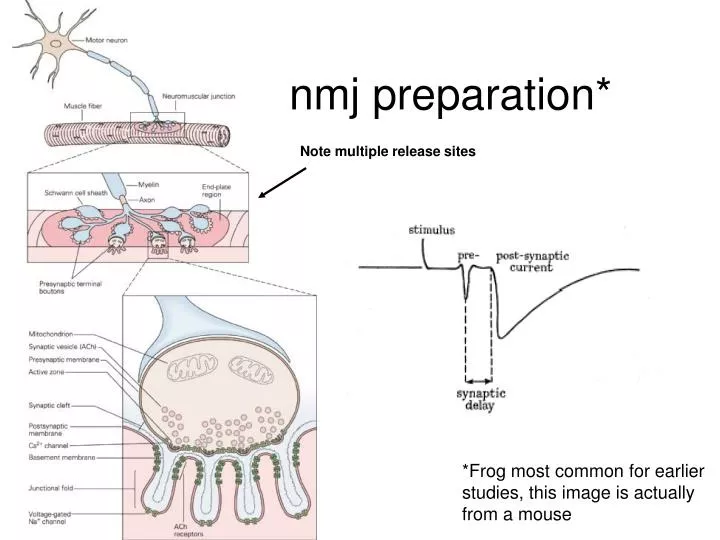
nmj preparation*. Note multiple release sites. *Frog most common for earlier studies, this image is actually from a mouse. Calcium is required for exocytosis. averaged responses. No Calcium. presynaptic action potential. Focal application of calcium. postsynaptic response—

E N D
nmj preparation* Note multiple release sites *Frog most common for earlier studies, this image is actually from a mouse
Calcium is required for exocytosis averaged responses No Calcium presynaptic action potential Focal application of calcium postsynaptic response— release of neurotransmitter Focal application of a little less calcium Back to no Calcium Frog neuromuscular junction Katz & Miledi 1965
The “calcium hypothesis”: Ca2+entry into the axon terminal rapidly triggers exocytosisby binding to a “calcium sensor” for release These experiments proved it Katz & Miledi 1967
At the Nernst Potential*, the balance of electrical and diffusion tendencies creates an electrochemical equilibrium between the opposing chemical (concentration) and electrical forces—NO NET MOVEMENT OF IONS Em = membrane potential R = Gas Constant T = Absolute Temperature z = valance of ion (charge) F = Faraday’s Constant At ~20, simplifies to: *AKA Reversal Potential, Equilibrium Potential
The amount of current carried by a particular ion across a membrane is a function of how many channels that are permeable to that ion are open (the “conductance”, or “g”), and the “driving force” for that ion, that is, how far the membrane potential is from the equilibrium potential for that ion V = I x R 1/R = g I = g x (Vm – Veq) For the calcium current: ICa = gCa x (Vm – ECa) ECa is very positive (>+100 mV), and gCa is voltage-dependent, so increasing membrane potential (depolarization) will open more and more calcium channels, increasing the conductance for calcium, but will also bring the membrane potential closer to ECa, reducing the driving force.
resting potential (membrane potential) calciumcurrent (U-shaped) g (increasing) driving force (decreasing)
“suppression” potential is approached what is this? note the growing “off response” what is this? full suppression is achieved during pulse Katz & Miledi 1967
Voltage-clamp recording of squid giant synapse Llinas, 1982
Release depends on [Ca]Ext 0.2 mM 0.25 mM more calcium = more release 0.3 mM Dodge & Rahamimoff, 1967
[Ca]/KCa n R ≈ 1 + [Ca]/KCa + [Mg]/KMg “Cooperativity” of the calcium dependence of release multiple calcium ion must bind to trigger release NOT linear n is typically estimated between 2 - 4 Dodge & Rahamimoff, 1967
How much calciuminside the cell is required forrelease from mammalian CNS neurons? -giant presynaptic terminal (Calyx of Held) is filled with “caged” calcium -a flash of light “uncages” the calcium; the brighter the flash, the more calcium is uncaged -a fluorescent calcium-indicator dye reports the concentration of calcium in the terminal -release of neurotransmitter is monitored as the postsynaptic response EPSCs Schneggenburger & Neher, 2000
Only a little bit of intracellular calcium (9-10 mM) is required to trigger release This is how much transmitter is released by an action potential Schneggenburger & Neher, 2000
C C room temp C C C D time How fast is release? fluorescent dyes Ipost current recording ICa Sabatini & Regehr, 1996 Very Fast less than 60 msec delay between start of ICa and Ipost at 38C
Calcium channels controlling release There are many different subtypes of voltage-gated calcium channels in neurons, including: L(onglasting)-, T(ransient)-, N(either)-, P(urkinje)-, Q(cool letter, in the right region of the alphabet)-, and R(esistant)-type channels …and this doesn’t even include ligand-gated calcium channels! How can we start to distinguish between them and identify their roles (if any) in controlling synaptic transmission?
Voltage-dependent calcium channels differ in their activation/inactivation ranges and kinetics (biophysical properties) T-Type note different time courses L-Type Miller, 1987
Calcium channels differ in their sensitivity to pharmacological reagents L-dihydropyridines (nifedipine, nimodipine, enhanced by BayK); rarely controls release N-1mM wConotoxin fraction GVIA (wCTx-GVIA) T-100 M Nickel (Ni2+); mostly localized to dendrites Q-1 M wAgatoxin fraction IVA (wAga-IVA) 1.5 M wConotoxin fraction MVIIC (wCTx-MVIIC) R-5 M wCTx-MVIIC P- 30 nM wAga-IVA All are blocked by 10 mM cobalt (Co2+) and cadmium (Cd2+) works pretty well, too.
(P-) (N-) (N-type) Relative response size (Q- & R-) time (min) We can use these tools to identify the channels that control neurotransmitter release Release at these hippocampal CA3-CA1 synapses is controlled by N-, and Q and(/or) R-type calcium channels; P-type channels do not control release at these synapses Wheeler et al, 1994
How many calcium channels does it take to release a vesicle? [Ca] “profile” release release no release Ca channel vesicle Mintz et al., 1995
Non-additive effects of toxins on release suggests that multiple calcium channels must open to trigger vesicle fusion at mammalian CNS synapses (note-higher conc. of wAga-IVA used here blocks P- and Q-type channels) more than 100% Mintz et al., 1995
Messages of the Day • Intracellular calcium triggers release of neurotransmitter • Calcium ions act cooperatively to trigger release (calcium-release relationship is not linear) • Very small increases in intracellular calcium (~10mM) trigger release very quickly (<100msec) • Different types of voltage-dependent calcium channels control release of neurotransmitter • Entry of calcium through more than one calcium channel may be required to trigger release
synaptic vesicle proteins Sudhof, 1995
{ SNARES and Synaptotagmin -synaptosomal-associated protein of 25 kDa (SNAP-25) -syntaxin -vesicle-associated membrane protein (VAMP aka synaptobrevin) Soluble NSF* Attachment Receptors *NEM-sensitive factor (NSF) (aka VAMP) Littleton et al., 2001
V-SNARES: Golgi/PM vacuole ER/golgi T-SNARES: plasma membrane vacuole Golgi/ER SNAREs were originally identified in yeast trafficking assays Only some combinations lead to fusion—leading to the hypothesis that SNAREs impart specificity to fusion reactions—and may be the “minimal machinery” required for vesicle fusion McNew et al., 2000
Tetanus toxin and various serotypes of Botulinum toxin cleave SNARE proteins: nb—tight form is insensitive to toxin Syntaxin is cleaved by BoNT/C SNAP-25 is cleaved by BoNT/A and E Vamp (synaptobrevin) is cleaved by TeNT, BoNT/B, D, F and G
Effects of toxins on exocytosis (spontaneous EPSCs) (vamp) (SNAP-25) (syntaxin) Capogna et al., 1997
Knocking out syntaxin abolishes all exocytosis TNT = Flies expressing TeTX throughout their nervous system to cleave VAMP (as with mice, some spontaneous release still present) Sys =Fly syntaxin knock-out Broadie et al., 1995
Knocking out Synaptotagmin 1 (SytI) abolishes the fast Ca2+-triggered component of exocytosis, but not the slower Ca2+-dependent “asynchronous” component of release asynchronous release remains in SytI KO asynchronous release Geppert et al., 1994
Synchronous vs. asynchronous release synchronous release asynchronous release 10 mM Ca2+ 10 mM Sr2+
Spontaneous mEPSCs are also unchanged in the SytI knock-out Wild-type 1.9 minis/synapse/min SytI KO 1.4 minis/synapse/min Geppert et al., 1994
Synaptotagmin I is required for fast Ca-triggered synaptic vesicle exocytosis, but not slower Ca-dependent asynchronous release or Ca-independent spontaneous release—could synaptotagmin I be a fast calcium sensor?
Synaptotagmin I: the calcium sensor? • Each C2 domain binds 3 Ca2+ ions w/affinities of 60 mM, 400 mM and >1 mM • C2A domain binds phospholipids in a Ca2+-dependent & cooperative manner • C2A domain binds syntaxin in a Ca2+-dependent manner (EC50=250 mM) • C2B domain mediates self-association of synaptotagmin I into multimers
EGTA Ca2+ GST Control Synaptotagmin Synaptotagmin I binds phospholipids (membranes) in the presence of calcium radiolabeled vesicle Syt heavy beads coupled to Syt are mixed with radioactive lipid vesicles, then spun down in a centrifuge, bringing along anything the Syt has bound to (unbound lipid vesicles stay in solution) Earles et al., 2001
Ca2+ binding to synaptotagmin I C2A domain: an “electrostatic switch”
= SNARE complex How could Synaptotagmin I control Ca-dependent exocytosis? Model II: Littleton et al., 2001 Model I: Shao et al., 1997 ) (Could this be a fusion pore? Either, neither, or both of these models could be correct
Messages of the Day • Presynaptic SNAREs are the minimal machinery required for both calcium-dependent and spontaneous fusion • Synaptotagmin is currently the best candidate for the fast Calcium Sensor triggering calcium-dependent synchronous release • A 2nd Calcium Sensor controls asynchronous release • The mechanisms underlying SNARE and synaptotagmin mediated fusion are hot topics of research—likely to be worked out within the next 5 years