Download

1 / 80
930 likes | 1.42k Views
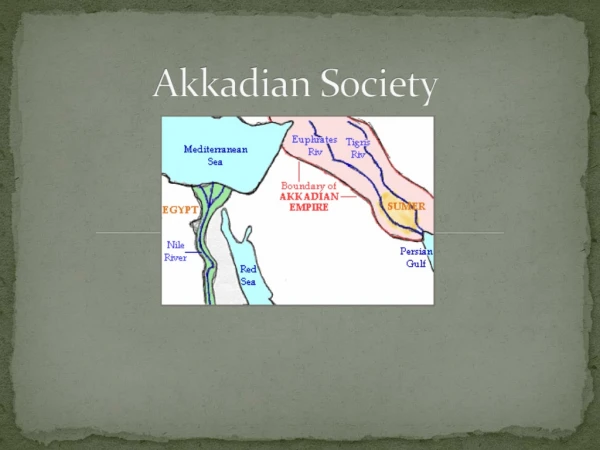
Evolution of sex determination and sex chromosomes : What can we learn from fish. What determines sex?. Sex determination : the process that makes the developmental decision whether the bipotential embryonic gonad analage starts to develop into an ovary or testes.

E N D
Evolution of sex determination and sex chromosomes : What can we learn from fish
What determines sex? Sex determination: the process that makes the developmental decision whether the bipotential embryonic gonad analage starts to develop into an ovary or testes Sex differentiation: the process that makes the functional ovary or testes from the undifferentiated gonad primordium, includes organogenesis, cell differentiation, meiosis Primary sex organs: gonads, germ cells (germline) + somatic cells Secondary sex characteristics: induced by hormones from the somatic cells of the gonad
The human Y: • - small, heterochromatic • - almost entirely excluded from meiosis • - degenerating • - gene-poor • is approx. 300 myrs old • - fully sequenced X Y
Modes of genetic sex determination I • Male heterogamety (XY-XX): • most mammals • -many amphibians (anurans and urodeles) • -some lizards • -many fish species (medaka) Female heterogamety (WZ-ZZ) -all birds -snakes -some lizards -many fish species (turbot, tilapia) -all (?) butterflies
Modes of genetic sex determination II Haplo-diploid systems: -haploid (unfertilized eggs) - male -diploid (fertilized eggs) - females (bees, ants) • X0 systems: • XX - female • -X0 - male • (grasshoppers) Autosome/X-chromosome ratio: -X:A=1 --- female -X:A=0.5 --- male (Drosophila, but: males have a Y-chromosome!)
Environmental sex determination -temperature: crocodilians: egg incubation temperature high - male low - female most turtles: egg incubation temperature high - female low - male -“environment“: Bonellia viridis dwarf males
Gonochorists Hermaphrodites Unisexuals Parthenogens Gynogens Hybridogens
Genetic sex determination + Environmental sex determination External influences can override the endogenous genetic sex determination: Environmental sex reversals
Coris julis Protogynoushermaphrodite
Amphiprion ocellatus Protandric hermaphrodite
Danio rerio Sex determination in zebrafish - no gonosomes identified so far (sex-linked traits?) - polygenic or environmental sex determination? - juvenile hermaphroditism (Takahashi 1977): all individuals first develop undifferentiated ovary-like gonads, oocyte apoptosis and spermatocyte differentation lead to testis development.
Tetraodon nigroviridis Sex determination in pufferfish: WZ/ZZ sex determination
Xiphophorus xiphidium - males are XY and females are XX - unbiased sex ratio, 1:1
Xiphophorus hellerii - polyfactorial genetic sex determination - several male (M) and female (F) determining factors, distributed over the whole genome - SM > SF determines male, SF > SM determines female de- velopment - strongly biased sex ratios
Xiphophorus alvarezi - Male are ZZ and females are WZ (Rio Candelaria population) - in the Rio San Ramon population autosomal modifiers exist
Xiphophorus nezahualcoyotl - most females are XX and all males are XY - occasionally XY females are obtained - different Y chromosomes exist, one (Y’) is susceptible to an autosomal modifier - the male sex determining locus on Y’ is blocked by the autosomal modifier
Xiphophorus multilineatus - all females are XX and most males (large and intermediate size) are XY - most small males are XX - an autosomal locus (A,a) affects sex differentiation - XX; AA fish become female, XX; aa become small males XX; Aa become predominantly female (20:1 female biased sex ratio)
Xiphophorus maculatus - three sex chromosomes coexist: W, X, Y - males are XY or YY, females are WY, WX or XX - rare autosomal modifiers cause XY females and XX males
female: XX male: XY
+277 females +75 males 7 8 0 1 3 M . . . . . 3 c 1 1 2 2 2 1 B 6 f 2 l L - 3 p c S s s p r Y Y a s e a C 1 k C Y r t a D r m S m d dmrt1bYmaps to the male sex determining locus females males cosmid 13 cosmid 9 control 1 2 3 4 5 6 1 2 3 4 5 6 [bp] dmrt1bY 965 casp6 1289
HNI (M-XY) SL1-dig (green) Markers from Linkage group 1 identify the sex chromosomes The gonosomes of medaka are homomorphic HNI(male, XY): SL1(green)
dmrt1bY (aka DMY) is the male sex determining gene of Medaka (Matsuda et al., Nature 2002, Nanda et al., PNAS 2002) The dmrt1a gene (LG9) has a duplicated copy, dmrt1bY, on the Y-chromosome X LG 9 LG 9 Y red: BAC 15 (dmrt1bY) green: SL1 (X and Y marker)
LG 1 LG 1 LG 9 LG 9 dmrt3b dmrt3b dmrt1b dmrt1bY transposons dmrt2 dmrt2 dmrt3a dmrt3 dmrt3 dmrt1a dmrt1 dmrt1 proto X Y proto Y X Duplication Insertion Degeneration/ Specialization Evolution of the Y-chromosome in medaka LG 1
X-chromosome flnk Y-chromosome 209O12 15H17 D011 168M02 113N21 95J10 125L10 Y specific region 20 kb KIAA0172 j MHCL D flnkR flnkL 10 kb j dmrt3 dmrt1bY transposon and repeat-sequence rich 260 kb 46kb autosome (LG 9) KIAA0172 MHCL dmrt1a dmrt3 dmrt2
- 4 Mya + -10 Mya + + (pseudogene) Dating of the gene duplication event of dmrt1 0.05 Phylogenetic tree (NJ) of Oryzias species from whole mitochondrial control region sequences Dating confirmed by substitution rates of nuclear genes
The medaka Y-chromosome is approximately 10 million years old The human Y is approximately 300 million years old
What can we learn from the medaka Y-chromosome about sex chromosome evolution? Prediction #1: Sex chromosomes evolve from (different) autosomes
7 8 0 1 3 M . . . . . 3 c 1 1 2 2 2 1 B 6 f 2 l L - 3 p c S s s p r Y Y a s e a C 1 k C Y r t a D r m S m d Genes outside the Y-specific region The duplicated fragment was inserted in a region with conserved syntenie to Hsa4 In stickleback the Y-chromosome shows conserved synteny with Hsa 15 In platyfish and guppy the sex determinining region shows conserved synteny with two other Hsas
What can we learn from the medaka Y-chromosome about sex chromosome evolution? Prediction #2: Sex chromosomes evolution starts with a stepwise allelic diversification of one (or two) loci on a pair of autosomes In Medaka sex chromosome evolution has started by a gene duplication process and no recombination at SD ab-inition
What can we learn from the medaka Y-chromosome about sex chromosome evolution? Prediction #3: Recombination around the sex determining gene is reduced
s t s O P Q 0 5 - 1 ( 0 ) Y c 1 ( 2 . 0 ) E R - 2 s t s O P Q 0 5 - 1 ( 0 ) s t E M 1 4 - 6 ( 1 3 . 8 ) Y c - 1 s t E M 1 4 - 6 s t E M 8 - 1 s t E M 3 2 - 5 s t E M 8 - 1 ( 1 5 . 8 ) s t E M 3 2 - 5 L ( 1 7 . 8 ) ( 0 ) s t E M 4 7 - 3 E R - 3 ( 1 8 . 4 ) ( 2 1 . 9 ) ( 3 . 3 ) ( 2 6 . 0 ) s t E M 4 7 - 3 ( 9 . 6 ) C 3 - 1 C 3 - 2 S L 2 d d 0 4 8 s t L C F - 2 E R - 2 s t E M 1 8 - 2 ( 2 1 . 0 ) E R - 3 ( 2 9 . 7 s t E M 8 - 8 ( 2 3 . 6 ) ( 3 5 . 6 ) E R - 1 ( 4 2 . 6 ) C 3 - 1 S L - 2 s t L C F - 2 d d 0 4 8 ( 3 0 . 1 ) s t E M 1 8 - 2 ( 5 3 . 3 ) ( 5 5 . 4 ) s t E M 8 - 8 E R - 1 ( 5 7 . 4 ) Genetic markers on the sex chromosomes of Medaka Sex reversed female meiosis Female meiosis Male meiosis X Y Y stsOPR04-1 Yc-2 y stsOPR04-1 Yc-2 Casp6 y Casp6 SL1 SL1 Casp3B Cen Casp3B Cen stsOPR04-1 lf Yc-2 Casp6 SL1 Cen lf Casp3B lf Sex chromosomal crossovers occur over almost the entire length of LG1: the pseudoautosomal region is very large but there is a small region of less than 3 cM of reduced recombination
What can we learn from the medaka Y-chromosome about sex chromosome evolution? Prediction #4: Due to reduced recombination genes degenerate
dmrt2 Duplicated genes found in the Y specific region dmrtbY dmrt3 j KIAA0172 MHCL MAP1 D Y-chromosome (BAC 113N21) dmrt1 dmrt3 KIAA0172 MHCL formin MAP1 autosomal dmrt1,3,2 cluster (LG9) autosomal MAP1 gene (LG18)
dmrt1bY is clearly a functional gene: - Spontaneous dmrt1bY mutants are XY male to female sex revertants - dmrt1bY BAC transgenics undergo XX female to male sex reversal - dmrt1bY is expressed in pre-somatic cells of the primordial gonad and in Sertoli cells .....and dmrt1bY appears to be the single gene involved
DMRT (Doublesex/Mab-3 Related Transcription factors) genes: • a multigene family in vertebrates (7-8 members in mammals), • dmrt1, 3, 2 are organized in a highly conserved cluster • - share the highly conserved DM domain, no similarity between • paralogs outside the DM domain • - putative transcription factors, intertwined zinc-fingers • - human DMRT 1 and 2 are putative dosage-dependent male sex • determination genes, which act downstream in the sex determination • cascade, haploinsufficiency is connected to XY male to female sex • reversal • - in birds Dmrt1 is located on the Z-chromosome
Sex determination/differentiation pathways: masters change, slaves remain Sry: eutherian mammals Sox9, WT1, SF1, AMH, (and possibly all other downstream components): vertebrates Dmrt1/Dsx/Mab3: vertebrates and invertebrates
Evolutionary fate of gene duplicates Degeneration Neofunctionalization Subfunctionalization
f m 5 Expression of dmrt1bY during embryogenesis fold expression 4,00 3,50 3,00 sex determination happens here: 2,50 2,00 1,50 1,00 0,50 1 2 3 4 5 6 f m f m f m f m f m f m f m f m days of development 7 8 11 13 14 20 30 40 dmrt1bY is expressed at gonad determination dmrt1a is not expressed before day 20 Real time RT-PCR
XX dmrt1bY transgenic XX female XY male dmrt1bY RNA in-situ histology 2 dph histology 30 dph Matsuda et al., PNAS, 104: 3865,2007
1400 1200 fold expression 1000 800 dmrt1a 60 eye gills skin eye gills skin liver liver brain brain heart ovary heart 40 kidney kidney spleen spleen muscle testes muscle male male female female 20 140 dmrt1bY 120 100 80 60 40 20 eye gills skin eye gills skin liver liver brain brain heart heart ovary kidney spleen kidney spleen testes muscle muscle Expression of dmrt1a and dmrt1bY in adults RNA in-situ hybridization: Sertoli cell specific expression Real time RT-PCR
Sequence comparison of the medaka dmrt1 promoters dmrt1bY dmrt1a
Repeat 3 sequences in the medaka genome The dmrt1bY promoter is derived from a novel class of transposable elements (P-like) “Izanaki“
Dmrt1bY transcriptional regulation The Britten-Davidson hypothesis (1971) Roy Britten Eric Davidson • evolutionary novelties require the rewiring of gene regulatory networks • dispersal of transposable elements allows the spread of regulatory motifs
Downregulation of the dmrt1bY promoter by Dmrt1bY and Dmrt1a Dmrt1
Electrophoretic Mobility Shift Analysis of in vitro translated Dmrt1bY and Dmrt1 protein wt competitor mutant competitor wt competitor mutant competitor + Dmrt1bY protein + Dmrt1a protein
Transcriptional rewiring of dmrt1bY by Izanaki shuts down dmrt1bY when it is not further needed and guarantees the survival of ist progenitor dmrt1a
9kb promoter::GFP stable transgenic line