Download
1 / 21
220 likes | 503 Views
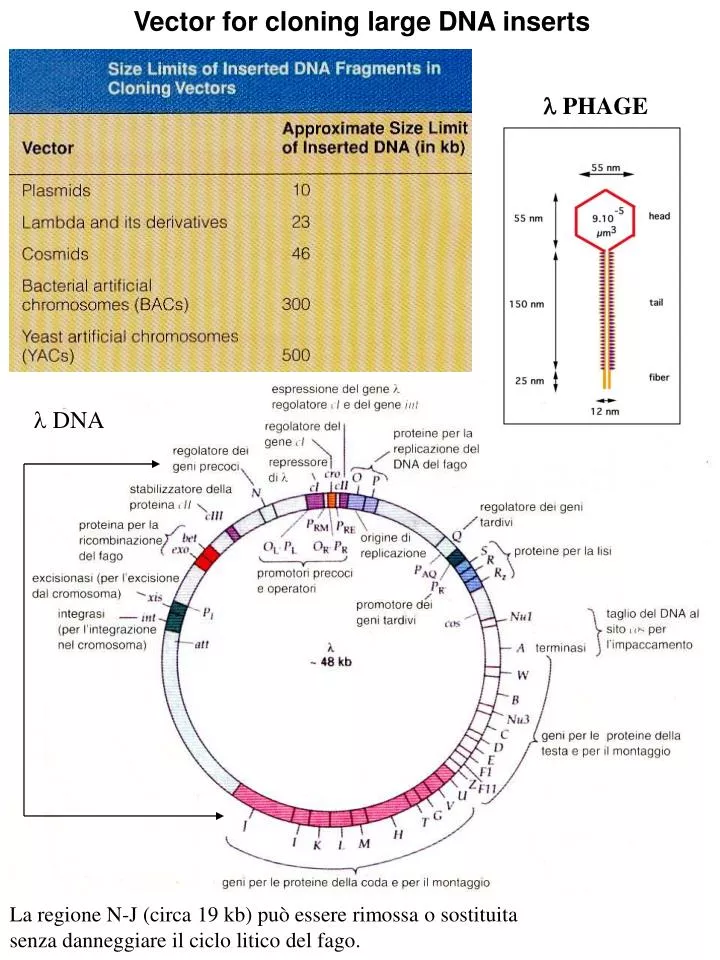
Vector for cloning large DNA inserts. l PHAGE. l DNA. La regione N-J (circa 19 kb) può essere rimossa o sostituita senza danneggiare il ciclo litico del fago. Replicazione del DNA del fago l durante il ciclo litico ed il ciclo lisogenico. Forma replicativa. Lambda phage vectors

E N D
Vector for cloning large DNA inserts l PHAGE l DNA La regione N-J (circa 19 kb) può essere rimossa o sostituita senza danneggiare il ciclo litico del fago.
Replicazione del DNA del fago l durante il ciclo litico ed il ciclo lisogenico Forma replicativa
Lambda phage vectors • Many l vector are constructed with a stufferfragment, a DNA segment that resides in the region between J and N genes. The stuffer fragment is cut out and replaced by foreign DNA during cloning procedure. • The stuffer fragment keeps the l vector at the correct size for packaging as a phage particle. • 2) The stuffer DNA fragment carries selectable marker genes that are removed when foreign DNA is inserted. About 60 % of lambda genome is needed for the lytic pathway
The lac5 gene encodes b-galactosidase allowing blue-white screening (cells lacZ- on medium containing IPTG and X-gal). EMBL series contain a polylinker with EcoRI, BamHI and SalI sites. Recombinant molecules are packaged into bacteriophage lambda heads in vitro. The product ofgene A cuts DNA at cos sites. The product of gene D is necessary for DNA packaging.
Impaccamento in vitro dei concatenameri di l in presenza della miscela dei due estratti di packaging Il processo di impaccamento in vitro si basa sulla miscelazione del DNA fagico ricombinante con elevate concentrazioni di precursori della testa, di code e proteine di packaging. Si utilizzano estratti concentrati di due particolari ceppi lisogeni per profagi l difettivi. Mutante nel gene D Mutante nel gene E Reazione di ligasi condotta in vitro Durante il processo di impaccamento i siti cos devono trovarsi ad un distanza compresa tra 37 e 52 kb. La proteina A taglia il DNA a livello dei siti cos.
Life cycle and DNA replication of M13 DNA cloning with single-stranded DNA vectors Sequencing of M13, f1 and fd filamentous coliphages indicates that they are very similar (~ 6400 base). Advantages of these vectors. Preparation of single-stranded DNA for sequencing (original deoxy method) and oligonucleotide directed mutagenesis. The RF can be purified and manipulated in vitro just like a plasmid. The RF is present at 100 copies per cell. For cloning purpose, DNA up to six times the length of M13can been packaged (~ 30 kb). Up to 1000 phages per cell To the M13 series of phage vectors belongs M13 mp, pEMBL, pBluescript, pSELECT. ori
Cosmid vector, a combination of λ and plasmid vectors Main features of cosmids (4-6 kb) 1)l cos sites. DNA fragments in the range of 30-46 kb cannot be cloned in lambda vectors but they can be cloned in cosmid vectors where large regions of l DNA are deleted. 2) a plasmid origin of replication 3) an antibiotic resistance 4) several unique restriction sites Foreign DNA is usuallydephosphorylated to prevent ligation of different fragments and/or circularization. Cut recombinant molecules at cos sites (product of gene A). The λ phage is needed only to inject the recombinant DNA into E. coli cells The cosmid contains the ori and so behaves as a plasmid replicating into E. coli cells. Cosmids are particularly attractive vectors for constructing libraries of eukaryotic genomes fragments. Possible products of the ligase reactions RE sites
Clonaggio mediante cosmidi Il fago l viene usato solo per inserire il DNA cosmidico all’interno della cellula di E. coli. Il cosmide viene tagliato in un sito BglII nei pressi di un sito cos. Il DNA genomico viene tagliato con Sau3A, enzima che produce estremità compatibili con quelle generate con BglII. Mescolando si ottiene una serie alternata inserto di DNA-vettore. Il fago viene impacchettato in vitro mediante un taglio a livello dei siti cos ad opera dell’endonucleasi A. Dopo l’inserimento nelle cellula batterica il cosmide ricircolarizza e si replica come un plasmide grazie alla presenza dell’origine di replicazione batterica (ori).
Cromosomi artificiali basati sul fago P1 (PACs) I PAC possono contenere fino a 100 kb. Dopo digestione con un enzima di restrizione (E) le estremità del PAC (braccio corto e lungo) vengono defosforilate. Ligasi del PAC con i frammenti da clonare. Taglio da parte dell’enzima Pacasi ed eliminazione del DNA in eccesso delle 115 kb. Impacchettamento in vitro del DNA in presenza delle teste e delle code. Le particelle fagiche mature iniettano il DNA in un ceppo di E. colicre+. E E La ricombinasi Cre agisce sui siti loxP producendo una molecola di plasmide P1 circolare (come λ) che viene mantenuto a basso numero di copie all’interno del batterio grazie all’origine di replicazione ori. Resistenza alla kanamicina. Per ottenere la produzione di un elevato numero di copie di P1 ricombinante è sufficiente indurre l’operone litico (repliconi P1) che è sotto il controllo del promotore lac. Normalmente il PAC è mantenuto in cellule che esprimono il repressore LacI
Bacterial Artificial Chromosomes (BACs) BACs (7 kb) are genetically engineered F factors that carry segment of foreign DNA as long as 300 kb. 1) oriS and repE control F-factor replication. 2) parA and parB genes limit the number of copies to 1 or 2 per cell. 3) A chloroamphenicol resistance gene. 4) A cloning segment including rare cutting enzymes as NotI and SfiI 5) BACs are introduced into E. coli host cells by electroporation. 6) BACs with PACs have been extensively used to construct human genomic DNA library.
Yeast Artificial Chromosomes (YACs) A non recombinant YAC is a circular plasmid that replicates in E. coli when it does not contain foreign DNA. YACs can accommodate up to 500 kb – 1 Mb DNA fragment. The foreignDNA is inserted YAC, into the EcoRI site and then the vector is linearized with BamHI. Then the YAC is transferred into yeast cells where it replicates as an eukaryotic chromosome. 1) ori is the plasmid origin of replication from pBR322 2) Ampr from pBR322 3) It contains the autonomous replication sequences, ARS, a yeast centromere sequence and 2 yeast telomere sequences (cloning site) BamHI EcoRI EcoRI BamHI
Cloning Strategies Shotgun cloning – Construction ofHuman genomic DNA library An EcoRI human genomic library may contain more than 700.000 different clones assuming that DNA fragments are 4 kb long (human haploid genome is 2,8x109 bp). For a cloned fragment of 20 kb the Minimum Number of Recombinants (MNR) is about 140.000 genome size = MNR Assuming the probability of including any sequence P = 95% and MNR = 140.000 (2,8x109: 20.000) the total number of recombinants (N) necessary for producing a representative gene library is 420.000 If P = 99%, N = 650.000 average length of cloned fragments ln (1-P) N= ln (1-1/MNR)
Detection of recombinant clones in a DNA library by colony hybridization
Probe identification and synthesis 1) heterologous probe 2) synthesis of a degenerate set of probes starting from the amino acid sequence of the protein 3) Single probes of 40-60 bases including inosine All 24 probes match this amino acids sequence: Trp-Asp-Arg-Asp-Met Met, Trp single codon Asp two codons Leu, Ser, Arg six codons Codon usage GUU, GUC, GUG and GUA are the 4 cognate codons for Val but in human 50 % of valine codons are GUG. 2 x 3 x 2 x 1 x 2 x 2 = 48 differenti oligonucleotidi 15-20 nucleotidi rappresentano un livello di specificità già sufficiente per trovare una sequenza unica all’interno di una genoteca.
cDNA cloning Una libreria di cDNA è più piccola (104-105 cloni) di una libreria genomica completa. Ad esempio nell’uomo abbiamo 30-35000 geni. Inoltre i frammenti di cDNA clonati hanno dimensioni ridotte (circa 2000 bp) rispetto a quelli comunemente usati per la costruzione di una libreria genomica (semplificazione delle procedure di clonaggio). Le librerie genomiche hanno il vantaggio di contenere i geni nella loro forma nativa che include le sequenze introniche e regioni regolative. Classica strategia per il clonaggio di cDNA Alternativamente a questo step possiamo inserire dei DNA linkers
Strategia alternativa per il clonaggio di cDNA che non prevede l’uso della nucleasi S1 ma della terminal trasferasi. L’enzima S1 è infatti relativamente poco puro e difficilmente controllabile, fattori che determinano il danneggiamento delle molecole di DNA. primer Primer-dG, DNA polimerasi (Klenow) + 4 dNTP Una libreria a cDNA permette l’espressione di proteine eucariotiche in cellule batteriche (i batteri non possono infatti rimuovere gli introni). Le librerie a cDNA sono usate per lo studio delle espressione genica (espressione tessuto-specifica e differenziamento). Le librerie a cDNA trovano un vasto impiego quando l’obiettivo finale è rappresentato dal prodotto proteico.
In vitro site-directed mutagenesis is an invaluable technique for studying: - protein structure-function relationships - gene expression - vector modification (insertion or deletion of restriction sites DNA cloning procedures) Protein engineering Regulation of transcription and translation -Mutant proteins facilitate the study of catalysis, substrate specificity, stability, etc. - Generation of chimeric proteins - Increasing the bioactivity of proteins (e.g. insulin, sequenase) - promoters - target sites for DNA binding proteins (activators, repressors) - translational apparatus components (mRNA, ribosomes, tRNA) Three different methods of site-directed mutagenesis are available: cassette mutagenesis primer extension procedures based on PCR Mutagenesi a cassetta Una breve sequenza sintetica di DNA contenente la mutazione desiderata viene inserita nel gene bersaglio al posto della corrispondente regione wild-type. • Svantaggi: • Siti di restrizione unici ai lati della regione da mutagenizzare. • Numero di oligonucleotidi da sintetizzare. Residui aminoacidici mutagenizzati N = A,T,G, C I = Inosina
Primer extension Single-stranded DNA vector (M13, pSELECT) The oligo pairing with the amp gene causes a four nucleotides insertion which repairs a frame shift mutation. Does not strand displace Repair deficient, suppress in vivo mismatch repair
Mutagenesi mediante PCR • Metodo del megaprimer • Vengono utilizzati tre primers e due reazioni di amplificazione. • Il prodotto della reazione di PCR viene utilizzato come primer (megaprimer) per la seconda reazione di amplificazione. Amplificazione con I° e II° primer (il prodotto rappresenta il megaprimer) Amplificazione con III° primer ed il megaprimer Un vantaggio è che la mutazione desiderata viene ottenuta con una frequenza praticamente del 100%. Uno svantaggio è invece legato alla bassa fedeltà replicativa della Taq polimerasi. Attualmente si usano enzimi quali Vent e Pfu dotati di attività 3'-5' exo (proofreading) e viene effettuato come controllo il sequenziamento dei prodotti di PCR.
Mutagenesi mediante PCR e DpnI I due primers mutagenici 5’-------TTC---------3’ 3’-------AAG---------5’ Il DNA stampo è metilato Questa tecnica di mutagenesi puo’ essere effettuata su qualsiasi vettore esi basa sul fatto che i prodotti di amplificazione non sono metilati. I primers portanti la mutazione sono di solito di 35-50 nt. Per la sintesi del DNA si usano enzimi quali Vent e Pfu dotati di attività 3'-5' exo (proofreading o correzione delle bozze) e si effettuano circa 18 cicli di PCR. DpnI digerisce sia il DNA metilato che quello emimetilato.
Sintesi di un gene artificiale samples (C mix) G A T C samples (C mix) Screening dei prodotti di mutagenesi (PCR-DpnI) mediante sequenziamento del DNA con il metodo di Sanger. Solo l’ultimo campione è uguale al wt (presenza di 2 C), gli altri sono tutti dei mutanti.